
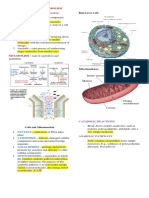
A Atividade Pif1 É Modulada Pela Sequência e Estrutura de DNA
A Atividade Pif1 É Modulada Pela Sequência e Estrutura de DNA
You might also like
- CH 18 Reading Guide - AnswersDocument6 pagesCH 18 Reading Guide - AnswersSylvia Grace0% (12)
- Mecanismo NADDocument12 pagesMecanismo NADlaura nataly sierra caicedoNo ratings yet
- Pnas 2207332119 (5478)Document12 pagesPnas 2207332119 (5478)rikoegawa25No ratings yet
- NIH Public Access: Author ManuscriptDocument21 pagesNIH Public Access: Author ManuscriptMariaNo ratings yet
- E STRUCTURATRIDIMENSIONALPEIADNDocument25 pagesE STRUCTURATRIDIMENSIONALPEIADNAngy ÑacatoNo ratings yet
- J. Biol. Chem.-2019-Chiu-2949-60Document13 pagesJ. Biol. Chem.-2019-Chiu-2949-60Dr. Kaushal Kishor SharmaNo ratings yet
- A Prebiotic Basis For ATP As The Universal Energy Currency - 2022Document25 pagesA Prebiotic Basis For ATP As The Universal Energy Currency - 2022cataawwwNo ratings yet
- Enzymology:: I Reveals Magnesium-Dependent Changes Kinetic Mechanism of Human DNA LigaseDocument10 pagesEnzymology:: I Reveals Magnesium-Dependent Changes Kinetic Mechanism of Human DNA LigaseVav VibavoNo ratings yet
- Elhafez Et Al. 2006Document12 pagesElhafez Et Al. 2006Chad SilbaNo ratings yet
- Transcription Factors / Overview 243Document9 pagesTranscription Factors / Overview 243Cindy GarciaNo ratings yet
- Cyclin PaperDocument14 pagesCyclin PapersayansoccercrazyNo ratings yet
- Into Thin Air How We Sense and Respond To HypoxiaDocument3 pagesInto Thin Air How We Sense and Respond To HypoxiaGabriela Catana TurcuNo ratings yet
- Mechanosensing of DNA Bending in A Single Specific protein-DNA ComplexDocument6 pagesMechanosensing of DNA Bending in A Single Specific protein-DNA ComplexLuis GonzálezNo ratings yet
- Asam Nukleat Dan Protein With SoundDocument18 pagesAsam Nukleat Dan Protein With SoundFika Nurul HafidzohNo ratings yet
- Cancer LetterDocument12 pagesCancer Letterwaqar AliNo ratings yet
- Midterm Biochem ReviewerDocument19 pagesMidterm Biochem ReviewerNATALIE NICOLE GABASNo ratings yet
- Gene Expression 2 Regulation: Slide 1Document20 pagesGene Expression 2 Regulation: Slide 1darkmagicdrakeNo ratings yet
- Genome-Wide Identification of RETINOBLASTOMA RELATED 1 Binding Sites in RegulatorsDocument35 pagesGenome-Wide Identification of RETINOBLASTOMA RELATED 1 Binding Sites in RegulatorsletsgopartaeNo ratings yet
- Apaf-1 AntibodyDocument1 pageApaf-1 AntibodySurendra Kumar KolliNo ratings yet
- Notes MolbioDocument8 pagesNotes Molbiochristineeraymundo02No ratings yet
- Caton Williams2011Document3 pagesCaton Williams2011Luis Carlos Vesga GamboaNo ratings yet
- C5RA10831DDocument21 pagesC5RA10831Dyook sungjaeNo ratings yet
- The Phosphate Makes A Difference Cellular Functions of NADPDocument10 pagesThe Phosphate Makes A Difference Cellular Functions of NADPYasif AbbasNo ratings yet
- 01 Atv 0000258979 9282Document7 pages01 Atv 0000258979 9282GeorgeNo ratings yet
- Molecular Basis of Inheritance NCERT HIGHLIGHT by SEEP PahujaDocument31 pagesMolecular Basis of Inheritance NCERT HIGHLIGHT by SEEP Pahujasafeerjavaid8No ratings yet
- A Comprehensive Evaluation of PCR Primers To Amplify The NifH Gene of NitrogenaseDocument12 pagesA Comprehensive Evaluation of PCR Primers To Amplify The NifH Gene of Nitrogenaseguy leviNo ratings yet
- Transduction Du Signal Redox-DépendantDocument3 pagesTransduction Du Signal Redox-DépendantMouad HiliaNo ratings yet
- Oxygen and Viruses: A Breathing Story: ReviewDocument4 pagesOxygen and Viruses: A Breathing Story: ReviewVedanthNo ratings yet
- Sublancin Is Not A Lantibiotic But An S-Linked - GlyDocument7 pagesSublancin Is Not A Lantibiotic But An S-Linked - Glyrusbianto wijayaNo ratings yet
- 16.blocking Peptides Against HBV - PreS1 Protein Selected From A Phage Display LibraryDocument5 pages16.blocking Peptides Against HBV - PreS1 Protein Selected From A Phage Display LibraryLeo SirsNo ratings yet
- SCIENCE 10 (4TH Quarter) - HeredityDocument7 pagesSCIENCE 10 (4TH Quarter) - HeredityJyña Khura TanoNo ratings yet
- European Journal of Medicinal ChemistryDocument12 pagesEuropean Journal of Medicinal ChemistryJafar HerizNo ratings yet
- Growth Phase DependentDocument7 pagesGrowth Phase DependentzaraNo ratings yet
- Journal Pgen 1010797Document35 pagesJournal Pgen 1010797jokkoNo ratings yet
- Central DogmaDocument4 pagesCentral DogmaalxndrasenalesNo ratings yet
- NF What's The Deal With Radicals?: KB - What Is It andDocument16 pagesNF What's The Deal With Radicals?: KB - What Is It andKost Vila SakinahNo ratings yet
- AP Biology Review SheetDocument4 pagesAP Biology Review Sheetapi-286357921No ratings yet
- Ch37 Riboflavinandbetaoxidationflavoenzymes GomesLabDocument23 pagesCh37 Riboflavinandbetaoxidationflavoenzymes GomesLabeka atika shafrianiNo ratings yet
- DNA Structure - PowerpointDocument29 pagesDNA Structure - PowerpointJohn Osborne100% (1)
- 179 FullDocument3 pages179 FullABDUL BASITNo ratings yet
- Computational Analysis of The Ligand Binding Site of The Extracellular ATP ReceptorDocument20 pagesComputational Analysis of The Ligand Binding Site of The Extracellular ATP ReceptorSaroj ManSingh BasnyatNo ratings yet
- Terraneo2014 PDFDocument10 pagesTerraneo2014 PDFYun AkbarNo ratings yet
- Module Ii-Genetics-G PDFDocument30 pagesModule Ii-Genetics-G PDFJennena Marie VeranoNo ratings yet
- From in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelDocument12 pagesFrom in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelAurora PradoNo ratings yet
- 1 s2.0 S1674205217301004 MainDocument15 pages1 s2.0 S1674205217301004 MainΚωνσταντίνος ΤσίπηςNo ratings yet
- Structure-Function Studies of DNA Polymerase LambdaDocument10 pagesStructure-Function Studies of DNA Polymerase LambdaGiovanni PiccinelliNo ratings yet
- Accepted Manuscript: Computational Biology and ChemistryDocument34 pagesAccepted Manuscript: Computational Biology and ChemistrynhkNo ratings yet
- Vegetarian AdaptationDocument4 pagesVegetarian AdaptationoanaraducuellarNo ratings yet
- Hypoxia-Inducible Factor 1: Regulation by Hypoxic and Non-Hypoxic ActivatorsDocument6 pagesHypoxia-Inducible Factor 1: Regulation by Hypoxic and Non-Hypoxic ActivatorsYun AkbarNo ratings yet
- Brain Sciences: Cofilin and Neurodegeneration: New Functions For An Old But Gold ProteinDocument12 pagesBrain Sciences: Cofilin and Neurodegeneration: New Functions For An Old But Gold Proteinjmarin55No ratings yet
- Spatiotemporal-Resolved Protein Networks Pro Filing With Photoactivation Dependent Proximity LabelingDocument12 pagesSpatiotemporal-Resolved Protein Networks Pro Filing With Photoactivation Dependent Proximity Labelingb00810902No ratings yet
- DPI O'DonnellDocument9 pagesDPI O'DonnellNadia DivakovaNo ratings yet
- J. Biol. Chem.-2003-Ayyagari-1618-25Document8 pagesJ. Biol. Chem.-2003-Ayyagari-1618-25A. M. Mahedi HasanNo ratings yet
- Regulação Metabolica 2013Document11 pagesRegulação Metabolica 2013Rafael GermanoNo ratings yet
- Assignment 2Document8 pagesAssignment 2Nesha VincentNo ratings yet
- Tunlid1994 PDFDocument9 pagesTunlid1994 PDFSuman KumarNo ratings yet
- TMP 8071Document6 pagesTMP 8071FrontiersNo ratings yet
- 1 s2.0 S2665928X19300054 MainDocument38 pages1 s2.0 S2665928X19300054 MainRocío Donaire CortésNo ratings yet
- NADPH OxidaseDocument9 pagesNADPH OxidaseAbhijeet MohantyNo ratings yet
- Natural Covalent Complexes of Nucleic Acids and Proteins: Some Comments On Practice and Theory On The Path From Well-Known Complexes To New OnesDocument6 pagesNatural Covalent Complexes of Nucleic Acids and Proteins: Some Comments On Practice and Theory On The Path From Well-Known Complexes To New OnesedeceNo ratings yet
- Cell Motility: From Molecules to OrganismsFrom EverandCell Motility: From Molecules to OrganismsAnne RidleyNo ratings yet
- A Enzima Moderadamente (D) Eficiente - Danos Relacionados À Catálise in Vivo e Seu ReparoDocument11 pagesA Enzima Moderadamente (D) Eficiente - Danos Relacionados À Catálise in Vivo e Seu ReparoRenan Guilherme de Oliveira GuihNo ratings yet
- Estrutura, Função e Termodinâmica de Lactato Desidrogenases de Humanos e o Parasita Da Malária P. FalciparumDocument14 pagesEstrutura, Função e Termodinâmica de Lactato Desidrogenases de Humanos e o Parasita Da Malária P. FalciparumRenan Guilherme de Oliveira GuihNo ratings yet
- Uma Abordagem Multifacetada de Busca de Acertos Revela Novos Ligantes Da Família LC3Document12 pagesUma Abordagem Multifacetada de Busca de Acertos Revela Novos Ligantes Da Família LC3Renan Guilherme de Oliveira GuihNo ratings yet
- Projeto de Uma Enzima Carregada Superpositivamente - Variante de Anidrase Carbônica Humana II Com Encapsulamento e Imobilização de FerritinaDocument14 pagesProjeto de Uma Enzima Carregada Superpositivamente - Variante de Anidrase Carbônica Humana II Com Encapsulamento e Imobilização de FerritinaRenan Guilherme de Oliveira GuihNo ratings yet
- Regulação Da Atividade Da MLL1 Metiltransferase em Dois Modos Distintos de Ligação Ao NucleossomoDocument9 pagesRegulação Da Atividade Da MLL1 Metiltransferase em Dois Modos Distintos de Ligação Ao NucleossomoRenan Guilherme de Oliveira GuihNo ratings yet
- Identificação de Uma Espécie Intermediária Ao Longo Da Via de Reação Da Nitrila Hidratase Por Espectroscopia EPRDocument12 pagesIdentificação de Uma Espécie Intermediária Ao Longo Da Via de Reação Da Nitrila Hidratase Por Espectroscopia EPRRenan Guilherme de Oliveira GuihNo ratings yet
- Para Elucidar o Eixo Microbiota-Cérebro Do Intestino Humano - Moléculas, Bioquímica e Implicações para A Saúde e DoençasDocument16 pagesPara Elucidar o Eixo Microbiota-Cérebro Do Intestino Humano - Moléculas, Bioquímica e Implicações para A Saúde e DoençasRenan Guilherme de Oliveira GuihNo ratings yet
- Caracterização Do Complexo Fuzzy de Alta Afinidade Entre o Domínio Desordenado Da Oncoproteína E7 Do HPV de Alto Risco e o Domínio TAZ2 Do CBPDocument12 pagesCaracterização Do Complexo Fuzzy de Alta Afinidade Entre o Domínio Desordenado Da Oncoproteína E7 Do HPV de Alto Risco e o Domínio TAZ2 Do CBPRenan Guilherme de Oliveira GuihNo ratings yet
- Investigação Estrutural e Bioquímica Da Ribonucleotídeo Redutase Classe I Do Hipertermófilo Aquifex AeolicusDocument15 pagesInvestigação Estrutural e Bioquímica Da Ribonucleotídeo Redutase Classe I Do Hipertermófilo Aquifex AeolicusRenan Guilherme de Oliveira GuihNo ratings yet
- Redução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoDocument5 pagesRedução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoRenan Guilherme de Oliveira GuihNo ratings yet
- Duplo Mutante Do Inibidor de Quimotripsina 2 Estabilizado Através Do Aumento Da Entropia ConformacionalDocument11 pagesDuplo Mutante Do Inibidor de Quimotripsina 2 Estabilizado Através Do Aumento Da Entropia ConformacionalRenan Guilherme de Oliveira GuihNo ratings yet
- Nearest-Neighbor E Ffects Modulate Loxp Spacer Dna Chemical Shifts and Guide Oligonucleotide Design For Nuclear Magnetic Resonance StudiesDocument10 pagesNearest-Neighbor E Ffects Modulate Loxp Spacer Dna Chemical Shifts and Guide Oligonucleotide Design For Nuclear Magnetic Resonance StudiesRenan Guilherme de Oliveira GuihNo ratings yet
- Mecanismos Bioquímicos Subjacentes À Neuroplasticidade Induzida Por PsicodélicosDocument10 pagesMecanismos Bioquímicos Subjacentes À Neuroplasticidade Induzida Por PsicodélicosRenan Guilherme de Oliveira GuihNo ratings yet
- Crescimento Dependente de Peptídeo em Levedura Através Da Expressão Genética Essencial Ativada Por Peptídeo FinoGPCRDocument10 pagesCrescimento Dependente de Peptídeo em Levedura Através Da Expressão Genética Essencial Ativada Por Peptídeo FinoGPCRRenan Guilherme de Oliveira GuihNo ratings yet
- Engenharia de Uma Variante de Endolisina Do Bacteriófago T7 Com Atividade de Amidase MelhoradaDocument15 pagesEngenharia de Uma Variante de Endolisina Do Bacteriófago T7 Com Atividade de Amidase MelhoradaRenan Guilherme de Oliveira GuihNo ratings yet
- Caracterização de Uma Enzima de Formação de Nitrogênio Envolvida Na Biossíntese de FosfazinomicinaDocument14 pagesCaracterização de Uma Enzima de Formação de Nitrogênio Envolvida Na Biossíntese de FosfazinomicinaRenan Guilherme de Oliveira GuihNo ratings yet
- Vidros Automobilísticos Como Vestígios de Cena de Crime - Uma Abordagem MultivariadaDocument8 pagesVidros Automobilísticos Como Vestígios de Cena de Crime - Uma Abordagem MultivariadaRenan Guilherme de Oliveira GuihNo ratings yet
- Identificação de Características Estruturais Distintas Do Domínio de Fusão de Proteína de Pico SARS-CoV-2 Essencial para Interação de MembranaDocument9 pagesIdentificação de Características Estruturais Distintas Do Domínio de Fusão de Proteína de Pico SARS-CoV-2 Essencial para Interação de MembranaRenan Guilherme de Oliveira GuihNo ratings yet
- CertificateDocument15 pagesCertificateAbhishek AryaNo ratings yet
- MutationDocument12 pagesMutationFlowerNo ratings yet
- GENE221 Progress Test RevisionDocument10 pagesGENE221 Progress Test RevisioncaryshealeyNo ratings yet
- Antibiotics 11 00125 v2Document13 pagesAntibiotics 11 00125 v2M Arfat YameenNo ratings yet
- Molecular Basis-WPS OfficeDocument33 pagesMolecular Basis-WPS OfficeShubhendu ChattopadhyayNo ratings yet
- Regulation of Histidine and Hut OperonsDocument11 pagesRegulation of Histidine and Hut Operonsaditi_joshee419No ratings yet
- Mmoi BC SbaDocument15 pagesMmoi BC SbaYoon YoonNo ratings yet
- Chapter 17 AnswersDocument15 pagesChapter 17 AnswersBen LucasNo ratings yet
- Cloning Tech Guide PDFDocument40 pagesCloning Tech Guide PDFLuis Fernando Soto UgaldiNo ratings yet
- Dna ReplicationDocument26 pagesDna ReplicationJerric Cristobal100% (3)
- Neuclic Acid and Protein SynthesisDocument21 pagesNeuclic Acid and Protein SynthesisPradyut BiswasNo ratings yet
- Unit 9 PDFDocument16 pagesUnit 9 PDFcarlette11No ratings yet
- 2020 F4 SCIENCE NOTES KSSM CHAPTER 4 6aDocument1 page2020 F4 SCIENCE NOTES KSSM CHAPTER 4 6aWani MesraNo ratings yet
- A Antisense Technology 1Document7 pagesA Antisense Technology 1Gourav JainNo ratings yet
- Quiz Prokaryotes Gene Regulation 1ST AttDocument4 pagesQuiz Prokaryotes Gene Regulation 1ST Attlasaten2350103No ratings yet
- Soal Biosel PDFDocument7 pagesSoal Biosel PDFjessicaNo ratings yet
- 2.7 DNA Replication, Transcription and TranslationDocument23 pages2.7 DNA Replication, Transcription and TranslationAleksandar KlingNo ratings yet
- Circular For IPFP Salary RevisionDocument6 pagesCircular For IPFP Salary RevisionAbdul RehmanNo ratings yet
- Bacterial ConjugationDocument39 pagesBacterial ConjugationIqra MujeebNo ratings yet
- 18 - 유전자의 발현 조절Document30 pages18 - 유전자의 발현 조절김도윤No ratings yet
- Plant PromotersDocument34 pagesPlant PromotersSreejesh P C0% (1)
- Central Dogma of LifeDocument2 pagesCentral Dogma of Lifepriya singhNo ratings yet
- Bac-to-Bac HBM Topo Secreted Expression System: User ManualDocument66 pagesBac-to-Bac HBM Topo Secreted Expression System: User ManualAhmad Bayquni BayquniNo ratings yet
- Temperate BacteriophageDocument26 pagesTemperate BacteriophageIzzah Mohd KhalilNo ratings yet
- Binary and Shuttle VectorDocument9 pagesBinary and Shuttle Vectordank memer100% (1)
- DNA Replication Sequencing Revision 2017Document58 pagesDNA Replication Sequencing Revision 2017Daniel sifuentes garciaNo ratings yet
- Shimshekgenesisi CRE2002Document9 pagesShimshekgenesisi CRE2002MihailNo ratings yet
- Blue WhiteDocument2 pagesBlue WhiteShyamala RamalingamNo ratings yet
- Alternative Splicing As A Source of Phenotypic DiversityDocument14 pagesAlternative Splicing As A Source of Phenotypic DiversitySergio VillicañaNo ratings yet









































































































Download as pdf or txt
You might also like
- CH 18 Reading Guide - AnswersDocument6 pagesCH 18 Reading Guide - AnswersSylvia Grace0% (12)
- Mecanismo NADDocument12 pagesMecanismo NADlaura nataly sierra caicedoNo ratings yet
- Pnas 2207332119 (5478)Document12 pagesPnas 2207332119 (5478)rikoegawa25No ratings yet
- NIH Public Access: Author ManuscriptDocument21 pagesNIH Public Access: Author ManuscriptMariaNo ratings yet
- E STRUCTURATRIDIMENSIONALPEIADNDocument25 pagesE STRUCTURATRIDIMENSIONALPEIADNAngy ÑacatoNo ratings yet
- J. Biol. Chem.-2019-Chiu-2949-60Document13 pagesJ. Biol. Chem.-2019-Chiu-2949-60Dr. Kaushal Kishor SharmaNo ratings yet
- A Prebiotic Basis For ATP As The Universal Energy Currency - 2022Document25 pagesA Prebiotic Basis For ATP As The Universal Energy Currency - 2022cataawwwNo ratings yet
- Enzymology:: I Reveals Magnesium-Dependent Changes Kinetic Mechanism of Human DNA LigaseDocument10 pagesEnzymology:: I Reveals Magnesium-Dependent Changes Kinetic Mechanism of Human DNA LigaseVav VibavoNo ratings yet
- Elhafez Et Al. 2006Document12 pagesElhafez Et Al. 2006Chad SilbaNo ratings yet
- Transcription Factors / Overview 243Document9 pagesTranscription Factors / Overview 243Cindy GarciaNo ratings yet
- Cyclin PaperDocument14 pagesCyclin PapersayansoccercrazyNo ratings yet
- Into Thin Air How We Sense and Respond To HypoxiaDocument3 pagesInto Thin Air How We Sense and Respond To HypoxiaGabriela Catana TurcuNo ratings yet
- Mechanosensing of DNA Bending in A Single Specific protein-DNA ComplexDocument6 pagesMechanosensing of DNA Bending in A Single Specific protein-DNA ComplexLuis GonzálezNo ratings yet
- Asam Nukleat Dan Protein With SoundDocument18 pagesAsam Nukleat Dan Protein With SoundFika Nurul HafidzohNo ratings yet
- Cancer LetterDocument12 pagesCancer Letterwaqar AliNo ratings yet
- Midterm Biochem ReviewerDocument19 pagesMidterm Biochem ReviewerNATALIE NICOLE GABASNo ratings yet
- Gene Expression 2 Regulation: Slide 1Document20 pagesGene Expression 2 Regulation: Slide 1darkmagicdrakeNo ratings yet
- Genome-Wide Identification of RETINOBLASTOMA RELATED 1 Binding Sites in RegulatorsDocument35 pagesGenome-Wide Identification of RETINOBLASTOMA RELATED 1 Binding Sites in RegulatorsletsgopartaeNo ratings yet
- Apaf-1 AntibodyDocument1 pageApaf-1 AntibodySurendra Kumar KolliNo ratings yet
- Notes MolbioDocument8 pagesNotes Molbiochristineeraymundo02No ratings yet
- Caton Williams2011Document3 pagesCaton Williams2011Luis Carlos Vesga GamboaNo ratings yet
- C5RA10831DDocument21 pagesC5RA10831Dyook sungjaeNo ratings yet
- The Phosphate Makes A Difference Cellular Functions of NADPDocument10 pagesThe Phosphate Makes A Difference Cellular Functions of NADPYasif AbbasNo ratings yet
- 01 Atv 0000258979 9282Document7 pages01 Atv 0000258979 9282GeorgeNo ratings yet
- Molecular Basis of Inheritance NCERT HIGHLIGHT by SEEP PahujaDocument31 pagesMolecular Basis of Inheritance NCERT HIGHLIGHT by SEEP Pahujasafeerjavaid8No ratings yet
- A Comprehensive Evaluation of PCR Primers To Amplify The NifH Gene of NitrogenaseDocument12 pagesA Comprehensive Evaluation of PCR Primers To Amplify The NifH Gene of Nitrogenaseguy leviNo ratings yet
- Transduction Du Signal Redox-DépendantDocument3 pagesTransduction Du Signal Redox-DépendantMouad HiliaNo ratings yet
- Oxygen and Viruses: A Breathing Story: ReviewDocument4 pagesOxygen and Viruses: A Breathing Story: ReviewVedanthNo ratings yet
- Sublancin Is Not A Lantibiotic But An S-Linked - GlyDocument7 pagesSublancin Is Not A Lantibiotic But An S-Linked - Glyrusbianto wijayaNo ratings yet
- 16.blocking Peptides Against HBV - PreS1 Protein Selected From A Phage Display LibraryDocument5 pages16.blocking Peptides Against HBV - PreS1 Protein Selected From A Phage Display LibraryLeo SirsNo ratings yet
- SCIENCE 10 (4TH Quarter) - HeredityDocument7 pagesSCIENCE 10 (4TH Quarter) - HeredityJyña Khura TanoNo ratings yet
- European Journal of Medicinal ChemistryDocument12 pagesEuropean Journal of Medicinal ChemistryJafar HerizNo ratings yet
- Growth Phase DependentDocument7 pagesGrowth Phase DependentzaraNo ratings yet
- Journal Pgen 1010797Document35 pagesJournal Pgen 1010797jokkoNo ratings yet
- Central DogmaDocument4 pagesCentral DogmaalxndrasenalesNo ratings yet
- NF What's The Deal With Radicals?: KB - What Is It andDocument16 pagesNF What's The Deal With Radicals?: KB - What Is It andKost Vila SakinahNo ratings yet
- AP Biology Review SheetDocument4 pagesAP Biology Review Sheetapi-286357921No ratings yet
- Ch37 Riboflavinandbetaoxidationflavoenzymes GomesLabDocument23 pagesCh37 Riboflavinandbetaoxidationflavoenzymes GomesLabeka atika shafrianiNo ratings yet
- DNA Structure - PowerpointDocument29 pagesDNA Structure - PowerpointJohn Osborne100% (1)
- 179 FullDocument3 pages179 FullABDUL BASITNo ratings yet
- Computational Analysis of The Ligand Binding Site of The Extracellular ATP ReceptorDocument20 pagesComputational Analysis of The Ligand Binding Site of The Extracellular ATP ReceptorSaroj ManSingh BasnyatNo ratings yet
- Terraneo2014 PDFDocument10 pagesTerraneo2014 PDFYun AkbarNo ratings yet
- Module Ii-Genetics-G PDFDocument30 pagesModule Ii-Genetics-G PDFJennena Marie VeranoNo ratings yet
- From in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelDocument12 pagesFrom in Silico To in Vitro: A Trip To Reveal Flavonoid Binding On The Rattus Norvegicus Kir6.1 ATP Sensitive Inward Rectifier Potassium ChannelAurora PradoNo ratings yet
- 1 s2.0 S1674205217301004 MainDocument15 pages1 s2.0 S1674205217301004 MainΚωνσταντίνος ΤσίπηςNo ratings yet
- Structure-Function Studies of DNA Polymerase LambdaDocument10 pagesStructure-Function Studies of DNA Polymerase LambdaGiovanni PiccinelliNo ratings yet
- Accepted Manuscript: Computational Biology and ChemistryDocument34 pagesAccepted Manuscript: Computational Biology and ChemistrynhkNo ratings yet
- Vegetarian AdaptationDocument4 pagesVegetarian AdaptationoanaraducuellarNo ratings yet
- Hypoxia-Inducible Factor 1: Regulation by Hypoxic and Non-Hypoxic ActivatorsDocument6 pagesHypoxia-Inducible Factor 1: Regulation by Hypoxic and Non-Hypoxic ActivatorsYun AkbarNo ratings yet
- Brain Sciences: Cofilin and Neurodegeneration: New Functions For An Old But Gold ProteinDocument12 pagesBrain Sciences: Cofilin and Neurodegeneration: New Functions For An Old But Gold Proteinjmarin55No ratings yet
- Spatiotemporal-Resolved Protein Networks Pro Filing With Photoactivation Dependent Proximity LabelingDocument12 pagesSpatiotemporal-Resolved Protein Networks Pro Filing With Photoactivation Dependent Proximity Labelingb00810902No ratings yet
- DPI O'DonnellDocument9 pagesDPI O'DonnellNadia DivakovaNo ratings yet
- J. Biol. Chem.-2003-Ayyagari-1618-25Document8 pagesJ. Biol. Chem.-2003-Ayyagari-1618-25A. M. Mahedi HasanNo ratings yet
- Regulação Metabolica 2013Document11 pagesRegulação Metabolica 2013Rafael GermanoNo ratings yet
- Assignment 2Document8 pagesAssignment 2Nesha VincentNo ratings yet
- Tunlid1994 PDFDocument9 pagesTunlid1994 PDFSuman KumarNo ratings yet
- TMP 8071Document6 pagesTMP 8071FrontiersNo ratings yet
- 1 s2.0 S2665928X19300054 MainDocument38 pages1 s2.0 S2665928X19300054 MainRocío Donaire CortésNo ratings yet
- NADPH OxidaseDocument9 pagesNADPH OxidaseAbhijeet MohantyNo ratings yet
- Natural Covalent Complexes of Nucleic Acids and Proteins: Some Comments On Practice and Theory On The Path From Well-Known Complexes To New OnesDocument6 pagesNatural Covalent Complexes of Nucleic Acids and Proteins: Some Comments On Practice and Theory On The Path From Well-Known Complexes To New OnesedeceNo ratings yet
- Cell Motility: From Molecules to OrganismsFrom EverandCell Motility: From Molecules to OrganismsAnne RidleyNo ratings yet
- A Enzima Moderadamente (D) Eficiente - Danos Relacionados À Catálise in Vivo e Seu ReparoDocument11 pagesA Enzima Moderadamente (D) Eficiente - Danos Relacionados À Catálise in Vivo e Seu ReparoRenan Guilherme de Oliveira GuihNo ratings yet
- Estrutura, Função e Termodinâmica de Lactato Desidrogenases de Humanos e o Parasita Da Malária P. FalciparumDocument14 pagesEstrutura, Função e Termodinâmica de Lactato Desidrogenases de Humanos e o Parasita Da Malária P. FalciparumRenan Guilherme de Oliveira GuihNo ratings yet
- Uma Abordagem Multifacetada de Busca de Acertos Revela Novos Ligantes Da Família LC3Document12 pagesUma Abordagem Multifacetada de Busca de Acertos Revela Novos Ligantes Da Família LC3Renan Guilherme de Oliveira GuihNo ratings yet
- Projeto de Uma Enzima Carregada Superpositivamente - Variante de Anidrase Carbônica Humana II Com Encapsulamento e Imobilização de FerritinaDocument14 pagesProjeto de Uma Enzima Carregada Superpositivamente - Variante de Anidrase Carbônica Humana II Com Encapsulamento e Imobilização de FerritinaRenan Guilherme de Oliveira GuihNo ratings yet
- Regulação Da Atividade Da MLL1 Metiltransferase em Dois Modos Distintos de Ligação Ao NucleossomoDocument9 pagesRegulação Da Atividade Da MLL1 Metiltransferase em Dois Modos Distintos de Ligação Ao NucleossomoRenan Guilherme de Oliveira GuihNo ratings yet
- Identificação de Uma Espécie Intermediária Ao Longo Da Via de Reação Da Nitrila Hidratase Por Espectroscopia EPRDocument12 pagesIdentificação de Uma Espécie Intermediária Ao Longo Da Via de Reação Da Nitrila Hidratase Por Espectroscopia EPRRenan Guilherme de Oliveira GuihNo ratings yet
- Para Elucidar o Eixo Microbiota-Cérebro Do Intestino Humano - Moléculas, Bioquímica e Implicações para A Saúde e DoençasDocument16 pagesPara Elucidar o Eixo Microbiota-Cérebro Do Intestino Humano - Moléculas, Bioquímica e Implicações para A Saúde e DoençasRenan Guilherme de Oliveira GuihNo ratings yet
- Caracterização Do Complexo Fuzzy de Alta Afinidade Entre o Domínio Desordenado Da Oncoproteína E7 Do HPV de Alto Risco e o Domínio TAZ2 Do CBPDocument12 pagesCaracterização Do Complexo Fuzzy de Alta Afinidade Entre o Domínio Desordenado Da Oncoproteína E7 Do HPV de Alto Risco e o Domínio TAZ2 Do CBPRenan Guilherme de Oliveira GuihNo ratings yet
- Investigação Estrutural e Bioquímica Da Ribonucleotídeo Redutase Classe I Do Hipertermófilo Aquifex AeolicusDocument15 pagesInvestigação Estrutural e Bioquímica Da Ribonucleotídeo Redutase Classe I Do Hipertermófilo Aquifex AeolicusRenan Guilherme de Oliveira GuihNo ratings yet
- Redução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoDocument5 pagesRedução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoRenan Guilherme de Oliveira GuihNo ratings yet
- Duplo Mutante Do Inibidor de Quimotripsina 2 Estabilizado Através Do Aumento Da Entropia ConformacionalDocument11 pagesDuplo Mutante Do Inibidor de Quimotripsina 2 Estabilizado Através Do Aumento Da Entropia ConformacionalRenan Guilherme de Oliveira GuihNo ratings yet
- Nearest-Neighbor E Ffects Modulate Loxp Spacer Dna Chemical Shifts and Guide Oligonucleotide Design For Nuclear Magnetic Resonance StudiesDocument10 pagesNearest-Neighbor E Ffects Modulate Loxp Spacer Dna Chemical Shifts and Guide Oligonucleotide Design For Nuclear Magnetic Resonance StudiesRenan Guilherme de Oliveira GuihNo ratings yet
- Mecanismos Bioquímicos Subjacentes À Neuroplasticidade Induzida Por PsicodélicosDocument10 pagesMecanismos Bioquímicos Subjacentes À Neuroplasticidade Induzida Por PsicodélicosRenan Guilherme de Oliveira GuihNo ratings yet
- Crescimento Dependente de Peptídeo em Levedura Através Da Expressão Genética Essencial Ativada Por Peptídeo FinoGPCRDocument10 pagesCrescimento Dependente de Peptídeo em Levedura Através Da Expressão Genética Essencial Ativada Por Peptídeo FinoGPCRRenan Guilherme de Oliveira GuihNo ratings yet
- Engenharia de Uma Variante de Endolisina Do Bacteriófago T7 Com Atividade de Amidase MelhoradaDocument15 pagesEngenharia de Uma Variante de Endolisina Do Bacteriófago T7 Com Atividade de Amidase MelhoradaRenan Guilherme de Oliveira GuihNo ratings yet
- Caracterização de Uma Enzima de Formação de Nitrogênio Envolvida Na Biossíntese de FosfazinomicinaDocument14 pagesCaracterização de Uma Enzima de Formação de Nitrogênio Envolvida Na Biossíntese de FosfazinomicinaRenan Guilherme de Oliveira GuihNo ratings yet
- Vidros Automobilísticos Como Vestígios de Cena de Crime - Uma Abordagem MultivariadaDocument8 pagesVidros Automobilísticos Como Vestígios de Cena de Crime - Uma Abordagem MultivariadaRenan Guilherme de Oliveira GuihNo ratings yet
- Identificação de Características Estruturais Distintas Do Domínio de Fusão de Proteína de Pico SARS-CoV-2 Essencial para Interação de MembranaDocument9 pagesIdentificação de Características Estruturais Distintas Do Domínio de Fusão de Proteína de Pico SARS-CoV-2 Essencial para Interação de MembranaRenan Guilherme de Oliveira GuihNo ratings yet
- CertificateDocument15 pagesCertificateAbhishek AryaNo ratings yet
- MutationDocument12 pagesMutationFlowerNo ratings yet
- GENE221 Progress Test RevisionDocument10 pagesGENE221 Progress Test RevisioncaryshealeyNo ratings yet
- Antibiotics 11 00125 v2Document13 pagesAntibiotics 11 00125 v2M Arfat YameenNo ratings yet
- Molecular Basis-WPS OfficeDocument33 pagesMolecular Basis-WPS OfficeShubhendu ChattopadhyayNo ratings yet
- Regulation of Histidine and Hut OperonsDocument11 pagesRegulation of Histidine and Hut Operonsaditi_joshee419No ratings yet
- Mmoi BC SbaDocument15 pagesMmoi BC SbaYoon YoonNo ratings yet
- Chapter 17 AnswersDocument15 pagesChapter 17 AnswersBen LucasNo ratings yet
- Cloning Tech Guide PDFDocument40 pagesCloning Tech Guide PDFLuis Fernando Soto UgaldiNo ratings yet
- Dna ReplicationDocument26 pagesDna ReplicationJerric Cristobal100% (3)
- Neuclic Acid and Protein SynthesisDocument21 pagesNeuclic Acid and Protein SynthesisPradyut BiswasNo ratings yet
- Unit 9 PDFDocument16 pagesUnit 9 PDFcarlette11No ratings yet
- 2020 F4 SCIENCE NOTES KSSM CHAPTER 4 6aDocument1 page2020 F4 SCIENCE NOTES KSSM CHAPTER 4 6aWani MesraNo ratings yet
- A Antisense Technology 1Document7 pagesA Antisense Technology 1Gourav JainNo ratings yet
- Quiz Prokaryotes Gene Regulation 1ST AttDocument4 pagesQuiz Prokaryotes Gene Regulation 1ST Attlasaten2350103No ratings yet
- Soal Biosel PDFDocument7 pagesSoal Biosel PDFjessicaNo ratings yet
- 2.7 DNA Replication, Transcription and TranslationDocument23 pages2.7 DNA Replication, Transcription and TranslationAleksandar KlingNo ratings yet
- Circular For IPFP Salary RevisionDocument6 pagesCircular For IPFP Salary RevisionAbdul RehmanNo ratings yet
- Bacterial ConjugationDocument39 pagesBacterial ConjugationIqra MujeebNo ratings yet
- 18 - 유전자의 발현 조절Document30 pages18 - 유전자의 발현 조절김도윤No ratings yet
- Plant PromotersDocument34 pagesPlant PromotersSreejesh P C0% (1)
- Central Dogma of LifeDocument2 pagesCentral Dogma of Lifepriya singhNo ratings yet
- Bac-to-Bac HBM Topo Secreted Expression System: User ManualDocument66 pagesBac-to-Bac HBM Topo Secreted Expression System: User ManualAhmad Bayquni BayquniNo ratings yet
- Temperate BacteriophageDocument26 pagesTemperate BacteriophageIzzah Mohd KhalilNo ratings yet
- Binary and Shuttle VectorDocument9 pagesBinary and Shuttle Vectordank memer100% (1)
- DNA Replication Sequencing Revision 2017Document58 pagesDNA Replication Sequencing Revision 2017Daniel sifuentes garciaNo ratings yet
- Shimshekgenesisi CRE2002Document9 pagesShimshekgenesisi CRE2002MihailNo ratings yet
- Blue WhiteDocument2 pagesBlue WhiteShyamala RamalingamNo ratings yet
- Alternative Splicing As A Source of Phenotypic DiversityDocument14 pagesAlternative Splicing As A Source of Phenotypic DiversitySergio VillicañaNo ratings yet