
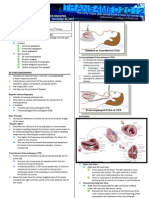
Blood Flow Mechanics in Cardiovascular Development
Blood Flow Mechanics in Cardiovascular Development
You might also like
- Bernstock, Classical MythologyDocument32 pagesBernstock, Classical MythologyNadia EvangelinaNo ratings yet
- Rosado-de-Christenson Localized Fibrous Tumors of The Pleura 2003Document25 pagesRosado-de-Christenson Localized Fibrous Tumors of The Pleura 2003JZNo ratings yet
- 23 Self-Sampling Blood Collection and Storage Device MarketDocument3 pages23 Self-Sampling Blood Collection and Storage Device MarketPavan GongaleNo ratings yet
- Disease/guide/diseases Cardiovascular#2Document10 pagesDisease/guide/diseases Cardiovascular#2Kristinelou Marie N. ReynaNo ratings yet
- Beam Modifying Devices in Radiotherapy.Document23 pagesBeam Modifying Devices in Radiotherapy.Pawan Kumar KandaraNo ratings yet
- Foldings of Embryo Foetal Period 1Document22 pagesFoldings of Embryo Foetal Period 1Riya SinghNo ratings yet
- CNS Part 2 Targeted NeurosonographyDocument11 pagesCNS Part 2 Targeted NeurosonographyHelena BravoNo ratings yet
- The Physiology of The Urine Production (2021)Document80 pagesThe Physiology of The Urine Production (2021)Shelly NobelNo ratings yet
- REVIEWCOURSEDocument102 pagesREVIEWCOURSEAjay KhadkaNo ratings yet
- Sept 2022 Belmont Poll Release - FinalDocument3 pagesSept 2022 Belmont Poll Release - FinalJack SterneNo ratings yet
- 6.04 - 6.06 UltrasoundDocument37 pages6.04 - 6.06 UltrasoundSneh KevadiyaNo ratings yet
- Thermal-Hydraulic Analysis of A Nuclear Research Reactor Core Channel PDFDocument12 pagesThermal-Hydraulic Analysis of A Nuclear Research Reactor Core Channel PDFAli AliNo ratings yet
- Ponciano IH PDFDocument69 pagesPonciano IH PDFCaressa Marie EstradaNo ratings yet
- Inflammatory Disease of BonesDocument93 pagesInflammatory Disease of BonesNelson LagatNo ratings yet
- LYMPHOGRAPHY-WPS OfficeDocument11 pagesLYMPHOGRAPHY-WPS OfficePax Giuseppe0% (1)
- Diagnostic Imaging - 11 - Regional Pathologies and Evaluation - Shoulder - Elbow - Wrist - Hand - Prof - Dr.i̇smet TAMERDocument34 pagesDiagnostic Imaging - 11 - Regional Pathologies and Evaluation - Shoulder - Elbow - Wrist - Hand - Prof - Dr.i̇smet TAMERAly MssreNo ratings yet
- CSFDocument21 pagesCSFjannatin aliya indrinaNo ratings yet
- Sorin Group s5 CatalogDocument36 pagesSorin Group s5 Cataloghadeel fahimNo ratings yet
- EU-5 EUS DiagnosisDocument28 pagesEU-5 EUS Diagnosiskeng suttisangchanNo ratings yet
- Memorandum OpinionDocument43 pagesMemorandum OpinionNEWS CENTER MaineNo ratings yet
- The Complete Guide To Vascular Ultrasound PDFDocument146 pagesThe Complete Guide To Vascular Ultrasound PDFAnna Sofía ParedesNo ratings yet
- Cross-Sectional Anatomy of The Thorax-BleinDocument355 pagesCross-Sectional Anatomy of The Thorax-BleinNatnael GetahunNo ratings yet
- Spectroscopy - B.Tech First YearDocument26 pagesSpectroscopy - B.Tech First YeartifinNo ratings yet
- SVT ShchukinDocument32 pagesSVT ShchukinСергей ЩукинNo ratings yet
- Mission1-General ECE Troubleshooting - LG-JRDocument32 pagesMission1-General ECE Troubleshooting - LG-JRKleber RodriguesNo ratings yet
- Intestinal ObstructionDocument15 pagesIntestinal ObstructionEmmeline Dycangchon-GarmaNo ratings yet
- Abnormalities of Fixation Saccade and Pursuit in Posterior Cortical AtropyDocument16 pagesAbnormalities of Fixation Saccade and Pursuit in Posterior Cortical AtropyDavid CristanchoNo ratings yet
- Upperlimb-Clinical CasesDocument18 pagesUpperlimb-Clinical CasesKambwe ChristopherNo ratings yet
- Long Term Care For Older Adults in Rural China Findings From The Recent Chinese Longitudinal Healthy Longevity SurveyDocument7 pagesLong Term Care For Older Adults in Rural China Findings From The Recent Chinese Longitudinal Healthy Longevity SurveyHerald Scholarly Open AccessNo ratings yet
- Mechanisms, Mitigation, and Management of Urinary Toxicity From Prostate RadiotherapyDocument10 pagesMechanisms, Mitigation, and Management of Urinary Toxicity From Prostate RadiotherapyZuriNo ratings yet
- Ancillary Studies in Pleural, Pericardial, and Peritoneal EffusionDocument9 pagesAncillary Studies in Pleural, Pericardial, and Peritoneal EffusionRoli CuizzNo ratings yet
- Osteoid Osteoma and Osteoblastoma: Review ArticleDocument12 pagesOsteoid Osteoma and Osteoblastoma: Review ArticleAnggi MartaNo ratings yet
- Overview of Colon Polyps - UpToDateDocument44 pagesOverview of Colon Polyps - UpToDateHartemes RosarioNo ratings yet
- Flow Cytometry: An Introduction and Application To Cytology: December 2018Document12 pagesFlow Cytometry: An Introduction and Application To Cytology: December 2018ABHINABA GUPTANo ratings yet
- Heresy Floorstanding SpeakerDocument1 pageHeresy Floorstanding Speaker96MysticNo ratings yet
- Cancer Treatment and Research CommunicationsDocument6 pagesCancer Treatment and Research CommunicationsCG MadeleineNo ratings yet
- Panoramic Radiographs: Technique & Anatomy Review: Continuing EducationDocument20 pagesPanoramic Radiographs: Technique & Anatomy Review: Continuing EducationRay AustraliaNo ratings yet
- Underwater Wireless-To-Plastic Optical Fiber Communication Systems With A Passive Front EndDocument3 pagesUnderwater Wireless-To-Plastic Optical Fiber Communication Systems With A Passive Front EndFilzaNo ratings yet
- Mammo Merged PDFDocument258 pagesMammo Merged PDFMartin AmoresNo ratings yet
- Biological Effects - Direct and Indirect - Trần Văn TiếnDocument14 pagesBiological Effects - Direct and Indirect - Trần Văn TiếnHgwergweg GwqegwNo ratings yet
- Drug Coasted Balloon Angioplasty Compared With Uncoated Balloons in The Treatment of Infrapopliteal Artery LesionsDocument7 pagesDrug Coasted Balloon Angioplasty Compared With Uncoated Balloons in The Treatment of Infrapopliteal Artery Lesionslia indria watiNo ratings yet
- Incisional HerniaDocument4 pagesIncisional HerniaRini YolandaNo ratings yet
- Node Place Analysis HubtropolisDocument9 pagesNode Place Analysis Hubtropolisrahel0% (1)
- eLORA Guidelines - DR Supplier - R1Document17 pageseLORA Guidelines - DR Supplier - R1Premananth ANo ratings yet
- NSG 461 - Disease Deep DiveDocument5 pagesNSG 461 - Disease Deep Diveapi-641353289100% (1)
- Color Doppler Ultrasound SysytemDocument4 pagesColor Doppler Ultrasound SysytemREJI RajuNo ratings yet
- Retrohepatic Gallbladder Masquerading As Hydatid Cyst in ADocument18 pagesRetrohepatic Gallbladder Masquerading As Hydatid Cyst in AMadalina BlagaNo ratings yet
- IBPJ Volume 20 Number 2 2021Document132 pagesIBPJ Volume 20 Number 2 2021Patricia Ferreira Dos SantosNo ratings yet
- 02 TissueDocument26 pages02 Tissuecow dasNo ratings yet
- Estimate Gadawara Power Plant NTPCDocument418 pagesEstimate Gadawara Power Plant NTPCSarin100% (1)
- Surgery FinalDocument70 pagesSurgery FinalIris BakerNo ratings yet
- 59 Minimally Invasive CV Surgery and ECMODocument7 pages59 Minimally Invasive CV Surgery and ECMOVictor PazNo ratings yet
- Lower Extremity RevascularizationDocument9 pagesLower Extremity RevascularizationleagagaNo ratings yet
- Cerebrospinal Fluid: Dr. Anupriya JDocument33 pagesCerebrospinal Fluid: Dr. Anupriya JVivek PatelNo ratings yet
- Scalp Block and Cervical Plexus Block Techniques - UpToDateDocument28 pagesScalp Block and Cervical Plexus Block Techniques - UpToDateAna Belén Artero CastañoNo ratings yet
- Breast MassDocument38 pagesBreast MassLester Paul SivilaNo ratings yet
- Radiolabelled Autologous Cells - Methods and Standardization For Clinical UseDocument130 pagesRadiolabelled Autologous Cells - Methods and Standardization For Clinical UseacbgdvNo ratings yet
- Walz 2015Document9 pagesWalz 2015Margarida ReisNo ratings yet
- Glenohumeral JointDocument85 pagesGlenohumeral JointPoojitha reddy MunnangiNo ratings yet
- Nexus ICSIDocument114 pagesNexus ICSIDr.Pratixa JoshiNo ratings yet
- Pathophysiology / Pharmacology: Cardiac PhysiologyDocument6 pagesPathophysiology / Pharmacology: Cardiac Physiologyjazzdoc007No ratings yet
- Prioritization of Nursing ProblemsDocument7 pagesPrioritization of Nursing ProblemsJoseph Raymund Fabian Huelar50% (4)
- Venous ReturnDocument2 pagesVenous ReturnBurnestNo ratings yet
- InggrisDocument13 pagesInggrisDefita DefitaNo ratings yet
- Non-Invasive Cardiac ImagingDocument9 pagesNon-Invasive Cardiac Imagingoddone_outNo ratings yet
- HortatoryDocument1 pageHortatoryDinda AmeliaNo ratings yet
- Coronary Angiography: A Continuing Education ProgramDocument71 pagesCoronary Angiography: A Continuing Education ProgramVincha R LuqmanNo ratings yet
- Cardiovascular EnduranceDocument7 pagesCardiovascular EnduranceFrancine ValdezNo ratings yet
- ECGInterpretation Cheat SheetDocument1 pageECGInterpretation Cheat SheetTom Mallinson100% (1)
- 2Document43 pages2myNo ratings yet
- Case Studies On Cardiovascular DiseasesDocument43 pagesCase Studies On Cardiovascular DiseasesClarise Barcenal BrionesNo ratings yet
- Self Asessment: Nama Karyawan Pendidikan Sertifikasi PenempatanDocument8 pagesSelf Asessment: Nama Karyawan Pendidikan Sertifikasi PenempatanRora Dhe OlalaNo ratings yet
- Circulatory Multiple Choice PDFDocument4 pagesCirculatory Multiple Choice PDFAndrea Jarani LinezoNo ratings yet
- Mrs. Maria Eloisa Lim Chu Gungon: When You Reach Age 75 When You Reach Age 75 If The Insured Passes Away at Age 75Document11 pagesMrs. Maria Eloisa Lim Chu Gungon: When You Reach Age 75 When You Reach Age 75 If The Insured Passes Away at Age 75John Philip TiongcoNo ratings yet
- General Biology 2: Quarter 4: Week 2 - Module 1B PDocument26 pagesGeneral Biology 2: Quarter 4: Week 2 - Module 1B PTipsy MonkeyNo ratings yet
- HOPE 1 Module 1 (Exercise For Fitness)Document10 pagesHOPE 1 Module 1 (Exercise For Fitness)Ma.Angelica B. AcedillaNo ratings yet
- ECG Minesota CodeDocument7 pagesECG Minesota CodeIman AzrbjNo ratings yet
- I. LESSON 5/melcs 4: Analyzes Physiological Indicators Such As Heart Rate, Rate ofDocument5 pagesI. LESSON 5/melcs 4: Analyzes Physiological Indicators Such As Heart Rate, Rate ofIssabhel Neyn DelgadoNo ratings yet
- History of Coronary ScoringDocument32 pagesHistory of Coronary ScoringAfiani JannahNo ratings yet
- Resurgence of Beta Blocker-Focus On HTDocument47 pagesResurgence of Beta Blocker-Focus On HTAkhil SharmaNo ratings yet
- Sinus Rhythm and Sinus and Atrial DysrhythmiasDocument41 pagesSinus Rhythm and Sinus and Atrial Dysrhythmiasclaudia brongNo ratings yet
- Cardiac Myocytes CultureDocument6 pagesCardiac Myocytes CultureÉmerson MoreiraNo ratings yet
- NCM 111 RRL Chapter 2Document5 pagesNCM 111 RRL Chapter 2kyle ibzNo ratings yet
- New England Journal Medicine: The ofDocument11 pagesNew England Journal Medicine: The ofahmadto80No ratings yet
- Genchembiochem Lab Report Compilation - 2Document40 pagesGenchembiochem Lab Report Compilation - 2Patrick PascuaNo ratings yet
- Regnier JujitsuDocument66 pagesRegnier JujitsuJesse Manek100% (1)
- Circulatory SystemDocument2 pagesCirculatory SystemcatchivanNo ratings yet
- Transport 1 MSDocument7 pagesTransport 1 MSAndrew KyawNo ratings yet
- ShockDocument30 pagesShockvinnu kalyanNo ratings yet
- Prasugrel and RosuvastatinDocument7 pagesPrasugrel and RosuvastatinMohammad Shahbaz AlamNo ratings yet























































































Download as pdf or txt
You might also like
- Bernstock, Classical MythologyDocument32 pagesBernstock, Classical MythologyNadia EvangelinaNo ratings yet
- Rosado-de-Christenson Localized Fibrous Tumors of The Pleura 2003Document25 pagesRosado-de-Christenson Localized Fibrous Tumors of The Pleura 2003JZNo ratings yet
- 23 Self-Sampling Blood Collection and Storage Device MarketDocument3 pages23 Self-Sampling Blood Collection and Storage Device MarketPavan GongaleNo ratings yet
- Disease/guide/diseases Cardiovascular#2Document10 pagesDisease/guide/diseases Cardiovascular#2Kristinelou Marie N. ReynaNo ratings yet
- Beam Modifying Devices in Radiotherapy.Document23 pagesBeam Modifying Devices in Radiotherapy.Pawan Kumar KandaraNo ratings yet
- Foldings of Embryo Foetal Period 1Document22 pagesFoldings of Embryo Foetal Period 1Riya SinghNo ratings yet
- CNS Part 2 Targeted NeurosonographyDocument11 pagesCNS Part 2 Targeted NeurosonographyHelena BravoNo ratings yet
- The Physiology of The Urine Production (2021)Document80 pagesThe Physiology of The Urine Production (2021)Shelly NobelNo ratings yet
- REVIEWCOURSEDocument102 pagesREVIEWCOURSEAjay KhadkaNo ratings yet
- Sept 2022 Belmont Poll Release - FinalDocument3 pagesSept 2022 Belmont Poll Release - FinalJack SterneNo ratings yet
- 6.04 - 6.06 UltrasoundDocument37 pages6.04 - 6.06 UltrasoundSneh KevadiyaNo ratings yet
- Thermal-Hydraulic Analysis of A Nuclear Research Reactor Core Channel PDFDocument12 pagesThermal-Hydraulic Analysis of A Nuclear Research Reactor Core Channel PDFAli AliNo ratings yet
- Ponciano IH PDFDocument69 pagesPonciano IH PDFCaressa Marie EstradaNo ratings yet
- Inflammatory Disease of BonesDocument93 pagesInflammatory Disease of BonesNelson LagatNo ratings yet
- LYMPHOGRAPHY-WPS OfficeDocument11 pagesLYMPHOGRAPHY-WPS OfficePax Giuseppe0% (1)
- Diagnostic Imaging - 11 - Regional Pathologies and Evaluation - Shoulder - Elbow - Wrist - Hand - Prof - Dr.i̇smet TAMERDocument34 pagesDiagnostic Imaging - 11 - Regional Pathologies and Evaluation - Shoulder - Elbow - Wrist - Hand - Prof - Dr.i̇smet TAMERAly MssreNo ratings yet
- CSFDocument21 pagesCSFjannatin aliya indrinaNo ratings yet
- Sorin Group s5 CatalogDocument36 pagesSorin Group s5 Cataloghadeel fahimNo ratings yet
- EU-5 EUS DiagnosisDocument28 pagesEU-5 EUS Diagnosiskeng suttisangchanNo ratings yet
- Memorandum OpinionDocument43 pagesMemorandum OpinionNEWS CENTER MaineNo ratings yet
- The Complete Guide To Vascular Ultrasound PDFDocument146 pagesThe Complete Guide To Vascular Ultrasound PDFAnna Sofía ParedesNo ratings yet
- Cross-Sectional Anatomy of The Thorax-BleinDocument355 pagesCross-Sectional Anatomy of The Thorax-BleinNatnael GetahunNo ratings yet
- Spectroscopy - B.Tech First YearDocument26 pagesSpectroscopy - B.Tech First YeartifinNo ratings yet
- SVT ShchukinDocument32 pagesSVT ShchukinСергей ЩукинNo ratings yet
- Mission1-General ECE Troubleshooting - LG-JRDocument32 pagesMission1-General ECE Troubleshooting - LG-JRKleber RodriguesNo ratings yet
- Intestinal ObstructionDocument15 pagesIntestinal ObstructionEmmeline Dycangchon-GarmaNo ratings yet
- Abnormalities of Fixation Saccade and Pursuit in Posterior Cortical AtropyDocument16 pagesAbnormalities of Fixation Saccade and Pursuit in Posterior Cortical AtropyDavid CristanchoNo ratings yet
- Upperlimb-Clinical CasesDocument18 pagesUpperlimb-Clinical CasesKambwe ChristopherNo ratings yet
- Long Term Care For Older Adults in Rural China Findings From The Recent Chinese Longitudinal Healthy Longevity SurveyDocument7 pagesLong Term Care For Older Adults in Rural China Findings From The Recent Chinese Longitudinal Healthy Longevity SurveyHerald Scholarly Open AccessNo ratings yet
- Mechanisms, Mitigation, and Management of Urinary Toxicity From Prostate RadiotherapyDocument10 pagesMechanisms, Mitigation, and Management of Urinary Toxicity From Prostate RadiotherapyZuriNo ratings yet
- Ancillary Studies in Pleural, Pericardial, and Peritoneal EffusionDocument9 pagesAncillary Studies in Pleural, Pericardial, and Peritoneal EffusionRoli CuizzNo ratings yet
- Osteoid Osteoma and Osteoblastoma: Review ArticleDocument12 pagesOsteoid Osteoma and Osteoblastoma: Review ArticleAnggi MartaNo ratings yet
- Overview of Colon Polyps - UpToDateDocument44 pagesOverview of Colon Polyps - UpToDateHartemes RosarioNo ratings yet
- Flow Cytometry: An Introduction and Application To Cytology: December 2018Document12 pagesFlow Cytometry: An Introduction and Application To Cytology: December 2018ABHINABA GUPTANo ratings yet
- Heresy Floorstanding SpeakerDocument1 pageHeresy Floorstanding Speaker96MysticNo ratings yet
- Cancer Treatment and Research CommunicationsDocument6 pagesCancer Treatment and Research CommunicationsCG MadeleineNo ratings yet
- Panoramic Radiographs: Technique & Anatomy Review: Continuing EducationDocument20 pagesPanoramic Radiographs: Technique & Anatomy Review: Continuing EducationRay AustraliaNo ratings yet
- Underwater Wireless-To-Plastic Optical Fiber Communication Systems With A Passive Front EndDocument3 pagesUnderwater Wireless-To-Plastic Optical Fiber Communication Systems With A Passive Front EndFilzaNo ratings yet
- Mammo Merged PDFDocument258 pagesMammo Merged PDFMartin AmoresNo ratings yet
- Biological Effects - Direct and Indirect - Trần Văn TiếnDocument14 pagesBiological Effects - Direct and Indirect - Trần Văn TiếnHgwergweg GwqegwNo ratings yet
- Drug Coasted Balloon Angioplasty Compared With Uncoated Balloons in The Treatment of Infrapopliteal Artery LesionsDocument7 pagesDrug Coasted Balloon Angioplasty Compared With Uncoated Balloons in The Treatment of Infrapopliteal Artery Lesionslia indria watiNo ratings yet
- Incisional HerniaDocument4 pagesIncisional HerniaRini YolandaNo ratings yet
- Node Place Analysis HubtropolisDocument9 pagesNode Place Analysis Hubtropolisrahel0% (1)
- eLORA Guidelines - DR Supplier - R1Document17 pageseLORA Guidelines - DR Supplier - R1Premananth ANo ratings yet
- NSG 461 - Disease Deep DiveDocument5 pagesNSG 461 - Disease Deep Diveapi-641353289100% (1)
- Color Doppler Ultrasound SysytemDocument4 pagesColor Doppler Ultrasound SysytemREJI RajuNo ratings yet
- Retrohepatic Gallbladder Masquerading As Hydatid Cyst in ADocument18 pagesRetrohepatic Gallbladder Masquerading As Hydatid Cyst in AMadalina BlagaNo ratings yet
- IBPJ Volume 20 Number 2 2021Document132 pagesIBPJ Volume 20 Number 2 2021Patricia Ferreira Dos SantosNo ratings yet
- 02 TissueDocument26 pages02 Tissuecow dasNo ratings yet
- Estimate Gadawara Power Plant NTPCDocument418 pagesEstimate Gadawara Power Plant NTPCSarin100% (1)
- Surgery FinalDocument70 pagesSurgery FinalIris BakerNo ratings yet
- 59 Minimally Invasive CV Surgery and ECMODocument7 pages59 Minimally Invasive CV Surgery and ECMOVictor PazNo ratings yet
- Lower Extremity RevascularizationDocument9 pagesLower Extremity RevascularizationleagagaNo ratings yet
- Cerebrospinal Fluid: Dr. Anupriya JDocument33 pagesCerebrospinal Fluid: Dr. Anupriya JVivek PatelNo ratings yet
- Scalp Block and Cervical Plexus Block Techniques - UpToDateDocument28 pagesScalp Block and Cervical Plexus Block Techniques - UpToDateAna Belén Artero CastañoNo ratings yet
- Breast MassDocument38 pagesBreast MassLester Paul SivilaNo ratings yet
- Radiolabelled Autologous Cells - Methods and Standardization For Clinical UseDocument130 pagesRadiolabelled Autologous Cells - Methods and Standardization For Clinical UseacbgdvNo ratings yet
- Walz 2015Document9 pagesWalz 2015Margarida ReisNo ratings yet
- Glenohumeral JointDocument85 pagesGlenohumeral JointPoojitha reddy MunnangiNo ratings yet
- Nexus ICSIDocument114 pagesNexus ICSIDr.Pratixa JoshiNo ratings yet
- Pathophysiology / Pharmacology: Cardiac PhysiologyDocument6 pagesPathophysiology / Pharmacology: Cardiac Physiologyjazzdoc007No ratings yet
- Prioritization of Nursing ProblemsDocument7 pagesPrioritization of Nursing ProblemsJoseph Raymund Fabian Huelar50% (4)
- Venous ReturnDocument2 pagesVenous ReturnBurnestNo ratings yet
- InggrisDocument13 pagesInggrisDefita DefitaNo ratings yet
- Non-Invasive Cardiac ImagingDocument9 pagesNon-Invasive Cardiac Imagingoddone_outNo ratings yet
- HortatoryDocument1 pageHortatoryDinda AmeliaNo ratings yet
- Coronary Angiography: A Continuing Education ProgramDocument71 pagesCoronary Angiography: A Continuing Education ProgramVincha R LuqmanNo ratings yet
- Cardiovascular EnduranceDocument7 pagesCardiovascular EnduranceFrancine ValdezNo ratings yet
- ECGInterpretation Cheat SheetDocument1 pageECGInterpretation Cheat SheetTom Mallinson100% (1)
- 2Document43 pages2myNo ratings yet
- Case Studies On Cardiovascular DiseasesDocument43 pagesCase Studies On Cardiovascular DiseasesClarise Barcenal BrionesNo ratings yet
- Self Asessment: Nama Karyawan Pendidikan Sertifikasi PenempatanDocument8 pagesSelf Asessment: Nama Karyawan Pendidikan Sertifikasi PenempatanRora Dhe OlalaNo ratings yet
- Circulatory Multiple Choice PDFDocument4 pagesCirculatory Multiple Choice PDFAndrea Jarani LinezoNo ratings yet
- Mrs. Maria Eloisa Lim Chu Gungon: When You Reach Age 75 When You Reach Age 75 If The Insured Passes Away at Age 75Document11 pagesMrs. Maria Eloisa Lim Chu Gungon: When You Reach Age 75 When You Reach Age 75 If The Insured Passes Away at Age 75John Philip TiongcoNo ratings yet
- General Biology 2: Quarter 4: Week 2 - Module 1B PDocument26 pagesGeneral Biology 2: Quarter 4: Week 2 - Module 1B PTipsy MonkeyNo ratings yet
- HOPE 1 Module 1 (Exercise For Fitness)Document10 pagesHOPE 1 Module 1 (Exercise For Fitness)Ma.Angelica B. AcedillaNo ratings yet
- ECG Minesota CodeDocument7 pagesECG Minesota CodeIman AzrbjNo ratings yet
- I. LESSON 5/melcs 4: Analyzes Physiological Indicators Such As Heart Rate, Rate ofDocument5 pagesI. LESSON 5/melcs 4: Analyzes Physiological Indicators Such As Heart Rate, Rate ofIssabhel Neyn DelgadoNo ratings yet
- History of Coronary ScoringDocument32 pagesHistory of Coronary ScoringAfiani JannahNo ratings yet
- Resurgence of Beta Blocker-Focus On HTDocument47 pagesResurgence of Beta Blocker-Focus On HTAkhil SharmaNo ratings yet
- Sinus Rhythm and Sinus and Atrial DysrhythmiasDocument41 pagesSinus Rhythm and Sinus and Atrial Dysrhythmiasclaudia brongNo ratings yet
- Cardiac Myocytes CultureDocument6 pagesCardiac Myocytes CultureÉmerson MoreiraNo ratings yet
- NCM 111 RRL Chapter 2Document5 pagesNCM 111 RRL Chapter 2kyle ibzNo ratings yet
- New England Journal Medicine: The ofDocument11 pagesNew England Journal Medicine: The ofahmadto80No ratings yet
- Genchembiochem Lab Report Compilation - 2Document40 pagesGenchembiochem Lab Report Compilation - 2Patrick PascuaNo ratings yet
- Regnier JujitsuDocument66 pagesRegnier JujitsuJesse Manek100% (1)
- Circulatory SystemDocument2 pagesCirculatory SystemcatchivanNo ratings yet
- Transport 1 MSDocument7 pagesTransport 1 MSAndrew KyawNo ratings yet
- ShockDocument30 pagesShockvinnu kalyanNo ratings yet
- Prasugrel and RosuvastatinDocument7 pagesPrasugrel and RosuvastatinMohammad Shahbaz AlamNo ratings yet