Download as docx, pdf, or txt
You might also like
- Biology Essay 1 - How Energy Is Transferred Within and Between OrganismsDocument3 pagesBiology Essay 1 - How Energy Is Transferred Within and Between OrganismsEva Dewsbury100% (3)
- Principles of Neurobiology 1st Edition Luo Test BankDocument15 pagesPrinciples of Neurobiology 1st Edition Luo Test BankOliviaHarrisonaxqyw100% (13)
- Biology: a QuickStudy Laminated Reference GuideFrom EverandBiology: a QuickStudy Laminated Reference GuideRating: 3 out of 5 stars3/5 (2)
- Achilike Sbi Assignment 1Document6 pagesAchilike Sbi Assignment 1skillsj123No ratings yet
- Biology Marking Scheme Paper I-Pre Joint 2015Document9 pagesBiology Marking Scheme Paper I-Pre Joint 2015josephNo ratings yet
- Photosynthesis and CRDocument5 pagesPhotosynthesis and CRKhyla Grace DijenoNo ratings yet
- Bioenergetics 20240416 114937 0000Document72 pagesBioenergetics 20240416 114937 0000Alexandra jade PagatpatanNo ratings yet
- Lecture I - Respiration and Photosynthesis Version 1Document14 pagesLecture I - Respiration and Photosynthesis Version 1Andre NesbethNo ratings yet
- Gen Bio1 ReviewerDocument4 pagesGen Bio1 ReviewerRhea SuarezNo ratings yet
- Finals Bio 1 RevDocument6 pagesFinals Bio 1 RevAnne Gelli ManaloNo ratings yet
- Cellular RespirationDocument18 pagesCellular RespirationConcepcion, Jemaelah P.No ratings yet
- Tun e Ni.Document9 pagesTun e Ni.LLORITO Cristell Joy M.No ratings yet
- Energy TransformationDocument14 pagesEnergy TransformationJoaquine ArateaNo ratings yet
- Cellular Respiration ExplanationDocument2 pagesCellular Respiration Explanationvestermail50% (2)
- Chapter 7 - Introduction To Cellular RespirationDocument14 pagesChapter 7 - Introduction To Cellular Respirationbabylen bahalaNo ratings yet
- Introduction To MetabolismDocument14 pagesIntroduction To MetabolismJohn Musa100% (1)
- GENERAL BIOLOGY Q2 WEEK 6 Glycolysis and Kreb CycleDocument14 pagesGENERAL BIOLOGY Q2 WEEK 6 Glycolysis and Kreb CycleAryan Jovic DomingoNo ratings yet
- Introduction To Cellular RespirationDocument5 pagesIntroduction To Cellular RespirationCarlton GrantNo ratings yet
- Measure Oxygen Uptake by RespirometerDocument17 pagesMeasure Oxygen Uptake by RespirometerTootsie100% (12)
- Reviewer in BiochemDocument11 pagesReviewer in BiochemLile RajabaNo ratings yet
- BIOTECHNOLOGY - Module 1,2,3Document24 pagesBIOTECHNOLOGY - Module 1,2,3CLIER GEROMALANo ratings yet
- Process of Cellular RespirationDocument3 pagesProcess of Cellular RespirationMercy NdungeNo ratings yet
- Cell RespirationDocument49 pagesCell RespirationJanine CambaNo ratings yet
- UntitledDocument14 pagesUntitledSyafrianda PanggabeanNo ratings yet
- BIOENERGETICSDocument3 pagesBIOENERGETICSJaemsNo ratings yet
- Midterm Review - BIODocument8 pagesMidterm Review - BIODolani N. AjanakuNo ratings yet
- Act. 6 Microbial PeraltaDocument9 pagesAct. 6 Microbial PeraltaCogie PeraltaNo ratings yet
- SLE111 Week 5 NotesDocument4 pagesSLE111 Week 5 NotesZahraNo ratings yet
- Biology Study Guide (Wheaton) Complete)Document7 pagesBiology Study Guide (Wheaton) Complete)Jireh HuangNo ratings yet
- Biology 2nd QRTR Exam - OutlineDocument5 pagesBiology 2nd QRTR Exam - OutlineLouie Joice MartinezNo ratings yet
- General Biology 101Document7 pagesGeneral Biology 101Czarina AsisNo ratings yet
- Ange EeeeeDocument4 pagesAnge EeeeeHazel BandayNo ratings yet
- Chloroplasts UpdatedDocument4 pagesChloroplasts Updateddrug123addict25No ratings yet
- Lesson 3 PDFDocument10 pagesLesson 3 PDFEdsa BaruaNo ratings yet
- Cemical Energy and ATP: ATP Is A Molecule With Three Phosphate Groups Attached To The EndDocument12 pagesCemical Energy and ATP: ATP Is A Molecule With Three Phosphate Groups Attached To The EndMhimi ViduyaNo ratings yet
- Cellular Respiration: Universidad Popular Del CesarDocument3 pagesCellular Respiration: Universidad Popular Del CesarNEFER PARRA TORRADONo ratings yet
- Photosynthesis and Cellular RespirationDocument8 pagesPhotosynthesis and Cellular RespirationFatima DariaNo ratings yet
- CHLOROPLASTDocument4 pagesCHLOROPLASTjyotirmoy choudhuryNo ratings yet
- Respiration in PlantsDocument6 pagesRespiration in PlantsAadil ShakulNo ratings yet
- Energy and RespirationDocument11 pagesEnergy and Respirationririrachma fitriahNo ratings yet
- Eals 11abma1 MangosingDocument34 pagesEals 11abma1 MangosingAndrea Mae V. MangosingNo ratings yet
- General Biology 1: Quarter 2 - Module 13 To 15: Energy TransformationDocument8 pagesGeneral Biology 1: Quarter 2 - Module 13 To 15: Energy Transformationkent ignacioNo ratings yet
- Þ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCDocument12 pagesÞ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCgauravNo ratings yet
- Bio I Master Guide - Test 2Document20 pagesBio I Master Guide - Test 2Brandon BarndtNo ratings yet
- How Cells Harvest EnergyDocument26 pagesHow Cells Harvest EnergyMarita YaghiNo ratings yet
- Concept of Cell Structures and FunctionsDocument3 pagesConcept of Cell Structures and FunctionsTyron ChuaNo ratings yet
- Photosynthesis ReadingDocument11 pagesPhotosynthesis ReadingLogan ParkisonNo ratings yet
- Elemental Composition of BacteriaDocument11 pagesElemental Composition of Bacteriaishmist04No ratings yet
- Bio 102 Concept ListDocument3 pagesBio 102 Concept ListLanae BraswellNo ratings yet
- Topic I: Cell: The Basic Unit of LifeDocument70 pagesTopic I: Cell: The Basic Unit of LifeJai CataluñaNo ratings yet
- Notes On Cellular RespirationDocument2 pagesNotes On Cellular Respirationaanonymousbros.0No ratings yet
- Life Sciences Grade 11 Energy and MetabolismDocument29 pagesLife Sciences Grade 11 Energy and MetabolismClaire WeerepasNo ratings yet
- BIOL 02 NotesDocument6 pagesBIOL 02 NotesAdam MoncktonNo ratings yet
- Name: Decemei M. Cuabo. Year & Section: Bs-Biology 1St Yr. 1ADocument3 pagesName: Decemei M. Cuabo. Year & Section: Bs-Biology 1St Yr. 1ADecemei CuaboNo ratings yet
- Biology Notes Unit TwoDocument158 pagesBiology Notes Unit TwoBlohsh KeenenNo ratings yet
- Cellular RespirationDocument10 pagesCellular RespirationXavier LecarosNo ratings yet
- Electron Transport ChainDocument4 pagesElectron Transport Chainavilashabas39No ratings yet
- Attempt1 - Final Biol101Document6 pagesAttempt1 - Final Biol101Mostafa A MansiNo ratings yet
- Reviewer in General Biology 1Document12 pagesReviewer in General Biology 1Christian Anthony SerquiñaNo ratings yet
- Carbohydrate Metabolism in Fish: o o o oDocument4 pagesCarbohydrate Metabolism in Fish: o o o oMD. SAKIB MUBALLIGNo ratings yet
- Energy TransformationDocument4 pagesEnergy TransformationAsh Nico BanquilloNo ratings yet
- Muscular and Skeletal Systems (Part 2)Document8 pagesMuscular and Skeletal Systems (Part 2)CynNo ratings yet
- Unit 1 - Markets in Action: Chapter 1 XDocument3 pagesUnit 1 - Markets in Action: Chapter 1 XCynNo ratings yet
- Account Formats and Statements LayoutsDocument5 pagesAccount Formats and Statements LayoutsCynNo ratings yet
- WEC11 - 01 - Que - 20190109 EcoDocument39 pagesWEC11 - 01 - Que - 20190109 EcoCynNo ratings yet
- The Effect of Profits or Losses On CapitalDocument2 pagesThe Effect of Profits or Losses On CapitalCynNo ratings yet
- Trading and Profit and Loss Accounts: Further ConsiderationsDocument3 pagesTrading and Profit and Loss Accounts: Further ConsiderationsCynNo ratings yet
- Sympathetic OphthalmiaDocument13 pagesSympathetic Ophthalmiamohamed salamaNo ratings yet
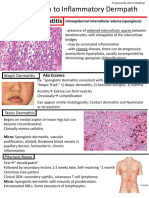
- Spongiotic Dermatitis: Aka EczemaDocument8 pagesSpongiotic Dermatitis: Aka EczemafadoNo ratings yet
- General Biology 1 Quarter 2 WEEK 1 Module 1bDocument15 pagesGeneral Biology 1 Quarter 2 WEEK 1 Module 1bNormal Fan100% (1)
- Psychology Module FinalsDocument36 pagesPsychology Module FinalsIrishNo ratings yet
- Kamota Mock Examinations 2023 s.6 BiologyDocument4 pagesKamota Mock Examinations 2023 s.6 Biologymungufeni amosNo ratings yet
- Agonist-Antagonist Paired Set Resistance Training. A Brief Review - Robbins Et Al. 2010Document10 pagesAgonist-Antagonist Paired Set Resistance Training. A Brief Review - Robbins Et Al. 2010Juan Andres Guzman WongNo ratings yet
- Dudwadkar 422017 AJMAH31948Document8 pagesDudwadkar 422017 AJMAH31948ROCHANA DUDWADKARNo ratings yet
- ANA PHY Lab Activity 6 Physiology of BonesDocument5 pagesANA PHY Lab Activity 6 Physiology of Bonesriya haryaniNo ratings yet
- AUBF LAB - Exams and QuizzesDocument18 pagesAUBF LAB - Exams and QuizzesLUALHATI VILLASNo ratings yet
- Theoharidesi, 2,3,4, S. Asadp, 5 and S. PanagiotidouiDocument7 pagesTheoharidesi, 2,3,4, S. Asadp, 5 and S. PanagiotidouiPsicoterapia InfantilNo ratings yet
- 2.5 EnzymesDocument42 pages2.5 EnzymesHelena GlanvilleNo ratings yet
- Prokaryotic Vs Eukaryotic Cells NotesDocument4 pagesProkaryotic Vs Eukaryotic Cells NotesMischi Jeda ElumbaNo ratings yet
- Advanced Science - 2020 - Teng - Shedding Light On Extracellular Vesicle Biogenesis and BioengineeringDocument17 pagesAdvanced Science - 2020 - Teng - Shedding Light On Extracellular Vesicle Biogenesis and BioengineeringNata1511No ratings yet
- Protos, Meaning of The First Rank' or of PrimeDocument16 pagesProtos, Meaning of The First Rank' or of Primepinkish7_preciousNo ratings yet
- FINAL CVS Module Time TableDocument5 pagesFINAL CVS Module Time TableAimen KhanNo ratings yet
- General Handout - ScienceDocument20 pagesGeneral Handout - ScienceRaven St. LouisNo ratings yet
- Legend PlexDocument12 pagesLegend Plexkike1790No ratings yet
- May 2021 Mark Scheme 2BDocument17 pagesMay 2021 Mark Scheme 2BMeenakshie ChaudrieNo ratings yet
- Africa by David Diop & Telephone Conversation by Wole SoyinkaDocument2 pagesAfrica by David Diop & Telephone Conversation by Wole Soyinkabaymax100% (2)
- Oogenesis and Ovarian Histology of The South American Lizard Salvator MerianaeDocument14 pagesOogenesis and Ovarian Histology of The South American Lizard Salvator MerianaeValeria Garcia ValdezNo ratings yet
- cc2861 PDFDocument6 pagescc2861 PDFArturo Eduardo Huarcaya OntiverosNo ratings yet
- Dr. Dinesh Kumar Yadav, KDCHDocument56 pagesDr. Dinesh Kumar Yadav, KDCHDinesh Kr. YadavNo ratings yet
- S6 Biology Paper 1 Set 1Document13 pagesS6 Biology Paper 1 Set 1Isaac Joshua goodNo ratings yet
- Wakefulness and SleepDocument76 pagesWakefulness and SleepXi En LookNo ratings yet
- Endocrinology Prepared By: Carl Leoneill Baroma, RMT, MPHDocument3 pagesEndocrinology Prepared By: Carl Leoneill Baroma, RMT, MPHCarl BaromaNo ratings yet
- Classification 2Document76 pagesClassification 2customer backup0% (1)
- Activity 4 Organogenesis in The Frog Embryo Activity Worksheet A. 4 MM Frog EmbryoDocument4 pagesActivity 4 Organogenesis in The Frog Embryo Activity Worksheet A. 4 MM Frog Embryojhayyypee jhayypsNo ratings yet
- SGLT2 Inhibitors and Mechanisms of Cardiovascular Benefit: A State-Of-The-Art ReviewDocument10 pagesSGLT2 Inhibitors and Mechanisms of Cardiovascular Benefit: A State-Of-The-Art ReviewLucian SiriteanuNo ratings yet
- Introduction To Human Anatomy and Physiology 4th Edition Solomon Test BankDocument10 pagesIntroduction To Human Anatomy and Physiology 4th Edition Solomon Test Bankmenaionemperilgtod26100% (28)