
Ferguson 2009
Ferguson 2009
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5824)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Hayward 2009Document13 pagesHayward 2009Ema DonNo ratings yet
- Ku Chen 2012Document6 pagesKu Chen 2012Ema DonNo ratings yet
- Nakamura 2012Document8 pagesNakamura 2012Ema DonNo ratings yet
- Strigo L Act OnesDocument7 pagesStrigo L Act OnesEma DonNo ratings yet
- Flower: Importance Morphology and Functions ClassificationsDocument10 pagesFlower: Importance Morphology and Functions ClassificationsEma DonNo ratings yet
- Main Functions of Roots:: and Produces Branch Roots Called Lateral RootsDocument4 pagesMain Functions of Roots:: and Produces Branch Roots Called Lateral RootsAleczandra QuesadaNo ratings yet
- Perk Buah Kemban H Mangg Pertum Ngan M GIS (Gar Mbuhan Morfolo Ndanp Ogi Dan Pemata N Fisiol Angan Logi AMADocument67 pagesPerk Buah Kemban H Mangg Pertum Ngan M GIS (Gar Mbuhan Morfolo Ndanp Ogi Dan Pemata N Fisiol Angan Logi AMAalvinaNo ratings yet
- RNS Price List 2019 FA WebDocument6 pagesRNS Price List 2019 FA WebnargueNo ratings yet
- Irish Sea Moss Supplements - Sea Moss by TracyDocument4 pagesIrish Sea Moss Supplements - Sea Moss by TracyTracy WNo ratings yet
- Kingdom PlantaeDocument17 pagesKingdom PlantaeHenry LimNo ratings yet
- Growth Characteristic of Welsh Onion Allium FistulDocument6 pagesGrowth Characteristic of Welsh Onion Allium FistulErick HidalgoNo ratings yet
- Active AbsorptionDocument2 pagesActive AbsorptionANo ratings yet
- Project List OrganicDocument50 pagesProject List OrganicSHAMPA SINHAROYNo ratings yet
- ASP Patch RecsDocument1 pageASP Patch RecsRochester EccNo ratings yet
- WWW Luxurydefinedcom April 2018pdfDocument2 pagesWWW Luxurydefinedcom April 2018pdfnudchaNo ratings yet
- Establishment of A Disease-Free Citrus Nursery System and Demonstration of Integrated Crop Health Management of Citrus OrchardsDocument6 pagesEstablishment of A Disease-Free Citrus Nursery System and Demonstration of Integrated Crop Health Management of Citrus OrchardsBrij Mohan SinghNo ratings yet
- 2018 - Effect of Integrated Nutrient Management and Different Plant Spacing On TulsiDocument10 pages2018 - Effect of Integrated Nutrient Management and Different Plant Spacing On TulsiNguyen Minh Tien B1509153No ratings yet
- Forest Biome EssayDocument2 pagesForest Biome EssayThesa TagalogNo ratings yet
- Soal Bahasa Inggris Adaptation of PlantsDocument2 pagesSoal Bahasa Inggris Adaptation of Plantsron noeNo ratings yet
- Sangay BangaloreDocument23 pagesSangay BangalorewinArunkumarNo ratings yet
- Amrut MittiDocument8 pagesAmrut Mittidonbosskiss100% (2)
- NematologyDocument46 pagesNematologyJayson BasiagNo ratings yet
- Cover Crops For Field Crop Systems: Fact Sheet 93 Agronomy Fact Sheet SeriesDocument2 pagesCover Crops For Field Crop Systems: Fact Sheet 93 Agronomy Fact Sheet SeriesGregory BakasNo ratings yet
- Video 1 - Functions of The Roots: Science ActivitiesDocument7 pagesVideo 1 - Functions of The Roots: Science ActivitiesGALYNSBELL ROJASNo ratings yet
- Actinorhizal Symbioses - Diversity and Bio GeographyDocument32 pagesActinorhizal Symbioses - Diversity and Bio Geographysaid_amrani7949No ratings yet
- TobaccoDocument91 pagesTobaccoFandy AchmadNo ratings yet
- Micropropagation of Self-Heading Philodendron Via Direct Shoot RegenerationDocument8 pagesMicropropagation of Self-Heading Philodendron Via Direct Shoot Regenerationmachaeallah4No ratings yet
- Reproduction PPT 3 ANGIOSPERMSDocument76 pagesReproduction PPT 3 ANGIOSPERMSlisanames.23No ratings yet
- A Taxonomic Revision of The Genus Aethionema S.L. (Brassicaceae) in IranDocument27 pagesA Taxonomic Revision of The Genus Aethionema S.L. (Brassicaceae) in IransalarNo ratings yet
- Week 08 Lab Prep - Plant BiologyDocument9 pagesWeek 08 Lab Prep - Plant BiologySyndyll OfianaNo ratings yet
- Bentley and Trimen-Medicinal Plants Vol 1 1875Document512 pagesBentley and Trimen-Medicinal Plants Vol 1 1875RonLayton100% (3)
- ASHITABADocument4 pagesASHITABATracy Gyneth Magno MedranoNo ratings yet
- Chapter 10 QuestionDocument21 pagesChapter 10 Questionapi-19650882No ratings yet
- Group DiscussionDocument4 pagesGroup DiscussionVijay RohillaNo ratings yet
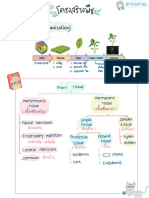
- สรุปโครงสร้างพืชDocument4 pagesสรุปโครงสร้างพืชรินบิNo ratings yet











































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5824)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Hayward 2009Document13 pagesHayward 2009Ema DonNo ratings yet
- Ku Chen 2012Document6 pagesKu Chen 2012Ema DonNo ratings yet
- Nakamura 2012Document8 pagesNakamura 2012Ema DonNo ratings yet
- Strigo L Act OnesDocument7 pagesStrigo L Act OnesEma DonNo ratings yet
- Flower: Importance Morphology and Functions ClassificationsDocument10 pagesFlower: Importance Morphology and Functions ClassificationsEma DonNo ratings yet
- Main Functions of Roots:: and Produces Branch Roots Called Lateral RootsDocument4 pagesMain Functions of Roots:: and Produces Branch Roots Called Lateral RootsAleczandra QuesadaNo ratings yet
- Perk Buah Kemban H Mangg Pertum Ngan M GIS (Gar Mbuhan Morfolo Ndanp Ogi Dan Pemata N Fisiol Angan Logi AMADocument67 pagesPerk Buah Kemban H Mangg Pertum Ngan M GIS (Gar Mbuhan Morfolo Ndanp Ogi Dan Pemata N Fisiol Angan Logi AMAalvinaNo ratings yet
- RNS Price List 2019 FA WebDocument6 pagesRNS Price List 2019 FA WebnargueNo ratings yet
- Irish Sea Moss Supplements - Sea Moss by TracyDocument4 pagesIrish Sea Moss Supplements - Sea Moss by TracyTracy WNo ratings yet
- Kingdom PlantaeDocument17 pagesKingdom PlantaeHenry LimNo ratings yet
- Growth Characteristic of Welsh Onion Allium FistulDocument6 pagesGrowth Characteristic of Welsh Onion Allium FistulErick HidalgoNo ratings yet
- Active AbsorptionDocument2 pagesActive AbsorptionANo ratings yet
- Project List OrganicDocument50 pagesProject List OrganicSHAMPA SINHAROYNo ratings yet
- ASP Patch RecsDocument1 pageASP Patch RecsRochester EccNo ratings yet
- WWW Luxurydefinedcom April 2018pdfDocument2 pagesWWW Luxurydefinedcom April 2018pdfnudchaNo ratings yet
- Establishment of A Disease-Free Citrus Nursery System and Demonstration of Integrated Crop Health Management of Citrus OrchardsDocument6 pagesEstablishment of A Disease-Free Citrus Nursery System and Demonstration of Integrated Crop Health Management of Citrus OrchardsBrij Mohan SinghNo ratings yet
- 2018 - Effect of Integrated Nutrient Management and Different Plant Spacing On TulsiDocument10 pages2018 - Effect of Integrated Nutrient Management and Different Plant Spacing On TulsiNguyen Minh Tien B1509153No ratings yet
- Forest Biome EssayDocument2 pagesForest Biome EssayThesa TagalogNo ratings yet
- Soal Bahasa Inggris Adaptation of PlantsDocument2 pagesSoal Bahasa Inggris Adaptation of Plantsron noeNo ratings yet
- Sangay BangaloreDocument23 pagesSangay BangalorewinArunkumarNo ratings yet
- Amrut MittiDocument8 pagesAmrut Mittidonbosskiss100% (2)
- NematologyDocument46 pagesNematologyJayson BasiagNo ratings yet
- Cover Crops For Field Crop Systems: Fact Sheet 93 Agronomy Fact Sheet SeriesDocument2 pagesCover Crops For Field Crop Systems: Fact Sheet 93 Agronomy Fact Sheet SeriesGregory BakasNo ratings yet
- Video 1 - Functions of The Roots: Science ActivitiesDocument7 pagesVideo 1 - Functions of The Roots: Science ActivitiesGALYNSBELL ROJASNo ratings yet
- Actinorhizal Symbioses - Diversity and Bio GeographyDocument32 pagesActinorhizal Symbioses - Diversity and Bio Geographysaid_amrani7949No ratings yet
- TobaccoDocument91 pagesTobaccoFandy AchmadNo ratings yet
- Micropropagation of Self-Heading Philodendron Via Direct Shoot RegenerationDocument8 pagesMicropropagation of Self-Heading Philodendron Via Direct Shoot Regenerationmachaeallah4No ratings yet
- Reproduction PPT 3 ANGIOSPERMSDocument76 pagesReproduction PPT 3 ANGIOSPERMSlisanames.23No ratings yet
- A Taxonomic Revision of The Genus Aethionema S.L. (Brassicaceae) in IranDocument27 pagesA Taxonomic Revision of The Genus Aethionema S.L. (Brassicaceae) in IransalarNo ratings yet
- Week 08 Lab Prep - Plant BiologyDocument9 pagesWeek 08 Lab Prep - Plant BiologySyndyll OfianaNo ratings yet
- Bentley and Trimen-Medicinal Plants Vol 1 1875Document512 pagesBentley and Trimen-Medicinal Plants Vol 1 1875RonLayton100% (3)
- ASHITABADocument4 pagesASHITABATracy Gyneth Magno MedranoNo ratings yet
- Chapter 10 QuestionDocument21 pagesChapter 10 Questionapi-19650882No ratings yet
- Group DiscussionDocument4 pagesGroup DiscussionVijay RohillaNo ratings yet
- สรุปโครงสร้างพืชDocument4 pagesสรุปโครงสร้างพืชรินบิNo ratings yet