
2012-Bacterium Lysinibacillus Sphaericus
2012-Bacterium Lysinibacillus Sphaericus
You might also like
- Herbal Bioactives and Food FortificationDocument274 pagesHerbal Bioactives and Food FortificationMilena Katarina StojiljkovicNo ratings yet
- CGH202008030750 - Lab M 2020 16384 - Laboratory - Covid PCR Test PDFDocument2 pagesCGH202008030750 - Lab M 2020 16384 - Laboratory - Covid PCR Test PDFMichael Vincent Naces AlmueteNo ratings yet
- Review CumarinasDocument39 pagesReview CumarinasLeidy GarciaNo ratings yet
- Recent Advances in Cordyceps Sinensis Polysaccharides Mycelial FermentationDocument16 pagesRecent Advances in Cordyceps Sinensis Polysaccharides Mycelial FermentationNguyễn Hữu ToànNo ratings yet
- Is Modulating Virus SIRDocument13 pagesIs Modulating Virus SIRThamer AlgohfailyNo ratings yet
- AntifungiDocument11 pagesAntifungiIdkNo ratings yet
- ForensikDocument19 pagesForensikKIMA 21019 Ferdianto sihotang SihotangNo ratings yet
- 2012-Bti Crystal ProteinDocument12 pages2012-Bti Crystal ProteinDwiNo ratings yet
- 2012-Bti Crystal ProteinDocument12 pages2012-Bti Crystal ProteinDwiNo ratings yet
- MónacoDocument23 pagesMónacopascoal shelzioNo ratings yet
- Drug Development Against Tuberculosis Impact of AlkaloidsDocument41 pagesDrug Development Against Tuberculosis Impact of Alkaloidsamine ben dahsenNo ratings yet
- Proc - Bichem2006 PatersonDocument8 pagesProc - Bichem2006 PatersonRashmi RamakrishnaNo ratings yet
- Aphid Resistance in Brassica Crops - Challenges, Biotechnological Progress and Emerging PossibilitiesDocument10 pagesAphid Resistance in Brassica Crops - Challenges, Biotechnological Progress and Emerging PossibilitiesVictor Lauro Perez GarciaNo ratings yet
- 2017-R-Pina-PErez-Antimicrobial Potential of Macro and Microalgae Against Pathogenic and Spoilage Microorganisms in FoodDocument11 pages2017-R-Pina-PErez-Antimicrobial Potential of Macro and Microalgae Against Pathogenic and Spoilage Microorganisms in FoodOscar Cabrera HerraizNo ratings yet
- !antimicrobials in BeekeepingDocument11 pages!antimicrobials in BeekeepingТетяна КозицькаNo ratings yet
- DIAREDocument21 pagesDIARETutorial Kelompok 5No ratings yet
- 2019 Book AdvancesInAnimalBiotechnologyDocument556 pages2019 Book AdvancesInAnimalBiotechnologypedro hernandezNo ratings yet
- 1 s2.0 S0169409X14000246 Main PDFDocument14 pages1 s2.0 S0169409X14000246 Main PDFCesar Rodolfo Angulo DelgadoNo ratings yet
- Beil Hartz 2017Document8 pagesBeil Hartz 2017andresleonardoNo ratings yet
- Química de Los AntibióticosDocument227 pagesQuímica de Los AntibióticosArturo García FloresNo ratings yet
- Goyal 2016Document29 pagesGoyal 2016Juan Krloz CastañedaNo ratings yet
- Verma 2007Document20 pagesVerma 2007jicama SEXTO NIVELNo ratings yet
- Industrial Crops and Products: Paulo R. Ribeiro, Renato D. de Castro, Luzimar G. FernandezDocument19 pagesIndustrial Crops and Products: Paulo R. Ribeiro, Renato D. de Castro, Luzimar G. FernandezCriztIan GgomesNo ratings yet
- Chrysobalanus Icaco - A Review of Its Phytochemistry and PharmacologyDocument7 pagesChrysobalanus Icaco - A Review of Its Phytochemistry and PharmacologyPatricia Oliveira VellanoNo ratings yet
- Phitoquímica y Farmacología IcacoDocument7 pagesPhitoquímica y Farmacología IcacoJhon GiraldoNo ratings yet
- Pharmaceutically Active Secondary Metabolites of Marine ActinobacteriaDocument17 pagesPharmaceutically Active Secondary Metabolites of Marine ActinobacteriakriukkoriukNo ratings yet
- Pharmacological Potential of Selected NaDocument10 pagesPharmacological Potential of Selected NaPedro Sánchez GarcíaNo ratings yet
- Ethnopharmacological and Phytochemical Attributes of Indian Tinospora Species: A Comprehensive ReviewDocument75 pagesEthnopharmacological and Phytochemical Attributes of Indian Tinospora Species: A Comprehensive ReviewSitti Rosdiana YahyaNo ratings yet
- 3 540 33526 9Document380 pages3 540 33526 9Rituporna NathNo ratings yet
- Nichols Etal 2008Document14 pagesNichols Etal 2008Julissa O CastilloNo ratings yet
- In Vitro and in Vivo Studies of Natural Products: A Challenge For Their Valuation. The Case Study of Chamomile (Matricaria Recutita L.)Document12 pagesIn Vitro and in Vivo Studies of Natural Products: A Challenge For Their Valuation. The Case Study of Chamomile (Matricaria Recutita L.)Eduardo SantiagoNo ratings yet
- Jamu (Ginger and Kaempferia) PDFDocument23 pagesJamu (Ginger and Kaempferia) PDFuntia_sariNo ratings yet
- PAK1 Blockers Potential Therapeutics Against CO 2020 Medicine in Drug DiscoDocument5 pagesPAK1 Blockers Potential Therapeutics Against CO 2020 Medicine in Drug DiscoKhánh TườngNo ratings yet
- Chemical Constituents of Bryophytes Bio and Chemical Diversity Biological Activity and Chemosystematics 1st Edition Yoshinori AsakawaDocument54 pagesChemical Constituents of Bryophytes Bio and Chemical Diversity Biological Activity and Chemosystematics 1st Edition Yoshinori Asakawajeannie.bookman296100% (18)
- Sanjai Saxena (Auth.) - Applied Microbiology-Springer India (2015)Document201 pagesSanjai Saxena (Auth.) - Applied Microbiology-Springer India (2015)Deepika KV100% (5)
- An Evaluation of Multipurpose Oil Seed Crop For Industrial Uses (Jatropha Curcas L.) : A ReviewDocument10 pagesAn Evaluation of Multipurpose Oil Seed Crop For Industrial Uses (Jatropha Curcas L.) : A ReviewYaru HageNo ratings yet
- ESBL in WildlifeDocument93 pagesESBL in WildlifevinodhpmdNo ratings yet
- Saussurea Lappa Review 2Document12 pagesSaussurea Lappa Review 2Mulayam Singh YadavNo ratings yet
- Lactobacillos RhamnosusDocument21 pagesLactobacillos RhamnosusDoris Banila San Martin OrellanaNo ratings yet
- Beneficial Features of Plant Growth-Promoting Rhizobacteria For Improving Plant Growth and Health in Challenging Conditions: A Methodical ReviewDocument21 pagesBeneficial Features of Plant Growth-Promoting Rhizobacteria For Improving Plant Growth and Health in Challenging Conditions: A Methodical ReviewagrocursoscafNo ratings yet
- Advanced Drug Delivery Reviews: Sarah S. Tang, Anucha Apisarnthanarak, Li Yang HsuDocument11 pagesAdvanced Drug Delivery Reviews: Sarah S. Tang, Anucha Apisarnthanarak, Li Yang HsuYariahna del Carmen Rossi MaussaNo ratings yet
- 0010 1A Book Angol 01 Novenyelettan PDFDocument121 pages0010 1A Book Angol 01 Novenyelettan PDFketakeeNo ratings yet
- Micotoxinas em AlimentosDocument12 pagesMicotoxinas em AlimentosTatiany RibeiroNo ratings yet
- Bacterial Associations With LegumesDocument27 pagesBacterial Associations With LegumesJorge CorderoNo ratings yet
- Plant Growth Promoting Bacterial EndophyDocument8 pagesPlant Growth Promoting Bacterial EndophyViníciusNo ratings yet
- Amanita PhalloidesDocument15 pagesAmanita PhalloidesJair Carrillo100% (1)
- Veterinary MycologyDocument194 pagesVeterinary MycologyMuhammad Noor Bumiputra100% (1)
- Mycotoxins and Their Effects On Human and Animal HealthDocument7 pagesMycotoxins and Their Effects On Human and Animal HealthtlatilpaNo ratings yet
- 2008.guava Review PaperDocument28 pages2008.guava Review PaperRomanauli SitumorangNo ratings yet
- 1 s2.0 S016816051530194X MainDocument17 pages1 s2.0 S016816051530194X MainverticalphoebusNo ratings yet
- (SpringerBriefs in Plant Science) Münir Öztürk, Volkan Altay, Khalid Rehman Hakeem, Eren Akçiçek - Liquorice-Springer International Publishing (2017)Document150 pages(SpringerBriefs in Plant Science) Münir Öztürk, Volkan Altay, Khalid Rehman Hakeem, Eren Akçiçek - Liquorice-Springer International Publishing (2017)Rosita HandayaniNo ratings yet
- Veterinary Microbiology: Ben Adler, Alejandro de La Pen A MoctezumaDocument10 pagesVeterinary Microbiology: Ben Adler, Alejandro de La Pen A MoctezumaNaufal WildanNo ratings yet
- Adler, Pen. Leptospira and Leptospirosis. 2010 PDFDocument10 pagesAdler, Pen. Leptospira and Leptospirosis. 2010 PDFWadramys SoumsNo ratings yet
- Bernard R. Glick (Auth.) - Beneficial Plant-Bacterial Interactions-Springer International Publishing (2015)Document251 pagesBernard R. Glick (Auth.) - Beneficial Plant-Bacterial Interactions-Springer International Publishing (2015)henrique.p.guimaraesNo ratings yet
- Forensic Mycology ReviewDocument11 pagesForensic Mycology ReviewDaniel MontealegreNo ratings yet
- Factors Affecting Mushroom GrowthDocument14 pagesFactors Affecting Mushroom Growthrommel5No ratings yet
- AmfDocument165 pagesAmfJesus OcampoNo ratings yet
- Arabidopsis Thaliana Legume Lectin-Like Proteins at The InterfaceDocument139 pagesArabidopsis Thaliana Legume Lectin-Like Proteins at The Interfaceكركشندي أفندي Karkshndy afndyNo ratings yet
- 1 s2.0 S1319562X21009918 MainDocument9 pages1 s2.0 S1319562X21009918 MainmerlinakisNo ratings yet
- Bioequivalence Studies in Drug Development: Methods and ApplicationsFrom EverandBioequivalence Studies in Drug Development: Methods and ApplicationsNo ratings yet
- A Transcriptomic Survey of TheDocument20 pagesA Transcriptomic Survey of TheDwiNo ratings yet
- Action of Natural PhytosanitarDocument10 pagesAction of Natural PhytosanitarDwiNo ratings yet
- 2020-Srikaya Seed Granule Different TemperatureDocument7 pages2020-Srikaya Seed Granule Different TemperatureDwiNo ratings yet
- 2017-Justicia Adhatoda MidgutDocument11 pages2017-Justicia Adhatoda MidgutDwiNo ratings yet
- 2014-Maybe Used-Ocotea CymbarumDocument10 pages2014-Maybe Used-Ocotea CymbarumDwiNo ratings yet
- A Toxin-Binding Alkaline PhospDocument20 pagesA Toxin-Binding Alkaline PhospDwiNo ratings yet
- 2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDocument7 pages2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDwiNo ratings yet
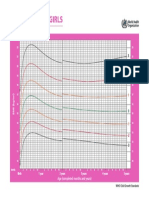
- CHT Bfa Girls Z 0 5Document1 pageCHT Bfa Girls Z 0 5DwiNo ratings yet
- A Transformed Bacterium ExpresDocument16 pagesA Transformed Bacterium ExpresDwiNo ratings yet
- A Transgenic Approach For ContDocument8 pagesA Transgenic Approach For ContDwiNo ratings yet
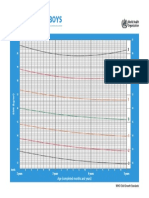
- CHT Bfa Boys Z 0 5Document1 pageCHT Bfa Boys Z 0 5DwiNo ratings yet
- CHT Bfa Boys Z 2 5Document1 pageCHT Bfa Boys Z 2 5DwiNo ratings yet
- Attachment 1706 631288cdc2e9fDocument4 pagesAttachment 1706 631288cdc2e9fDwiNo ratings yet
- 2012-Eichornia Crasiper-CulexDocument7 pages2012-Eichornia Crasiper-CulexDwiNo ratings yet
- Attachment 1706 631288cdc271dDocument5 pagesAttachment 1706 631288cdc271dDwiNo ratings yet
- 2012-Fungitoxic InsecticidalDocument18 pages2012-Fungitoxic InsecticidalDwiNo ratings yet
- 2012-Garlic-Allium Sativum EndotoxinDocument11 pages2012-Garlic-Allium Sativum EndotoxinDwiNo ratings yet
- 2012-CYP450 On EarthDocument12 pages2012-CYP450 On EarthDwiNo ratings yet
- Aedes Aegypti Bacillus ThuringiensisDocument9 pagesAedes Aegypti Bacillus ThuringiensisDwiNo ratings yet
- 2012-Crimson Speckled MothDocument10 pages2012-Crimson Speckled MothDwiNo ratings yet
- 2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDocument682 pages2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDwiNo ratings yet
- 2012 Citrus Sinensis Musca DomesticaDocument8 pages2012 Citrus Sinensis Musca DomesticaDwiNo ratings yet
- 2012-Castor Bean Leaf-Spodoptera LituraDocument7 pages2012-Castor Bean Leaf-Spodoptera LituraDwiNo ratings yet
- 2012-Bti-Applied and Environmental MicrobiologyDocument6 pages2012-Bti-Applied and Environmental MicrobiologyDwiNo ratings yet
- 2012-Bti Residual TMOFDocument5 pages2012-Bti Residual TMOFDwiNo ratings yet
- 2012-Calatropis GiganteaDocument14 pages2012-Calatropis GiganteaDwiNo ratings yet
- 2012-Bti SV2, Serratia Nematodipila SV6Document7 pages2012-Bti SV2, Serratia Nematodipila SV6DwiNo ratings yet
- LESSON 1 Explain The Postulates of The Cell TheoryDocument28 pagesLESSON 1 Explain The Postulates of The Cell Theorysophia DelacruzNo ratings yet
- Science Magazine - 30 June 2023Document136 pagesScience Magazine - 30 June 2023Pablo GarcíaNo ratings yet
- Biology Notes PDF 1 Beq2woDocument373 pagesBiology Notes PDF 1 Beq2woarmanmalik5216No ratings yet
- Disorders and Diseases of Immune SystemDocument21 pagesDisorders and Diseases of Immune SystemWijesiri D WNo ratings yet
- Biomolecules WorksheetDocument4 pagesBiomolecules WorksheetJana ArafaNo ratings yet
- Mtle - Blood Banking (Revised)Document25 pagesMtle - Blood Banking (Revised)docv526No ratings yet
- How To Construct A Nature Summary Paragraph: Show" or Their Equivalent)Document1 pageHow To Construct A Nature Summary Paragraph: Show" or Their Equivalent)31 Roihan FauziNo ratings yet
- Special Staining: Intan Chairun Nisa, S.Si., M.BiotechDocument46 pagesSpecial Staining: Intan Chairun Nisa, S.Si., M.BiotechKecil Dek100% (1)
- V.O.T.E. Mock Test 1: "Champions Keep Playing Until They Get It Right."Document45 pagesV.O.T.E. Mock Test 1: "Champions Keep Playing Until They Get It Right."Miten GandhiNo ratings yet
- Genetic Disorders ProjectDocument2 pagesGenetic Disorders ProjectbarkhaNo ratings yet
- Hemifacial Microsomia: Presented by NithishDocument29 pagesHemifacial Microsomia: Presented by NithishNithish ReddyNo ratings yet
- Microbial Structures and Functions MoicroorganismsDocument390 pagesMicrobial Structures and Functions MoicroorganismssamsoatsNo ratings yet
- Impact of GMOsDocument4 pagesImpact of GMOstony montanNo ratings yet
- Dairy Cattle Breeding ManagementDocument34 pagesDairy Cattle Breeding Managementaginche amareNo ratings yet
- ANAT 2033 EoR Test 1 2015Document6 pagesANAT 2033 EoR Test 1 2015Matsiri ImmanuelNo ratings yet
- Chapter 7 Protoplast CultureDocument76 pagesChapter 7 Protoplast Culturerajiv pathakNo ratings yet
- WBC PathoDocument29 pagesWBC PathoMadhumitha ElangovanNo ratings yet
- Combining Ability Estimates in Virus Resistant and Susceptible Lines of ChilliDocument5 pagesCombining Ability Estimates in Virus Resistant and Susceptible Lines of ChilliShailendra RajanNo ratings yet
- Student Copy of GeneticsDocument122 pagesStudent Copy of GeneticsJofran TisuelaNo ratings yet
- Biology f4t2 Pp3 MsDocument4 pagesBiology f4t2 Pp3 MssamuwelNo ratings yet
- Evaluation For Module 1Document4 pagesEvaluation For Module 1Patricia PascuaNo ratings yet
- Motivational LetterDocument1 pageMotivational LetterZubair GhouriNo ratings yet
- Msczo 501Document213 pagesMsczo 501vvaishali784No ratings yet
- Tahiri (2022)Document22 pagesTahiri (2022)Jorge Andres Molina GonzalezNo ratings yet
- محاضرات ضبط الجودة أ.د. أحمد عبد المولىDocument79 pagesمحاضرات ضبط الجودة أ.د. أحمد عبد المولىMohammed AhmedNo ratings yet
- DNA Evidence in Contemporary Legal Perspective: Institutional ArticleDocument7 pagesDNA Evidence in Contemporary Legal Perspective: Institutional ArticleVijaykumar D SNo ratings yet
- 0610 BIOLOGY: MARK SCHEME For The October/November 2015 SeriesDocument14 pages0610 BIOLOGY: MARK SCHEME For The October/November 2015 SeriesKashviNo ratings yet
- Origin of Life: Earth Evolving UniverseDocument1 pageOrigin of Life: Earth Evolving UniversemateothgodNo ratings yet
- Unit 1.2 Mendelian GeneticsDocument7 pagesUnit 1.2 Mendelian GeneticsRegine Coeli Menta LansanganNo ratings yet


















































































































Download as pdf or txt
You might also like
- Herbal Bioactives and Food FortificationDocument274 pagesHerbal Bioactives and Food FortificationMilena Katarina StojiljkovicNo ratings yet
- CGH202008030750 - Lab M 2020 16384 - Laboratory - Covid PCR Test PDFDocument2 pagesCGH202008030750 - Lab M 2020 16384 - Laboratory - Covid PCR Test PDFMichael Vincent Naces AlmueteNo ratings yet
- Review CumarinasDocument39 pagesReview CumarinasLeidy GarciaNo ratings yet
- Recent Advances in Cordyceps Sinensis Polysaccharides Mycelial FermentationDocument16 pagesRecent Advances in Cordyceps Sinensis Polysaccharides Mycelial FermentationNguyễn Hữu ToànNo ratings yet
- Is Modulating Virus SIRDocument13 pagesIs Modulating Virus SIRThamer AlgohfailyNo ratings yet
- AntifungiDocument11 pagesAntifungiIdkNo ratings yet
- ForensikDocument19 pagesForensikKIMA 21019 Ferdianto sihotang SihotangNo ratings yet
- 2012-Bti Crystal ProteinDocument12 pages2012-Bti Crystal ProteinDwiNo ratings yet
- 2012-Bti Crystal ProteinDocument12 pages2012-Bti Crystal ProteinDwiNo ratings yet
- MónacoDocument23 pagesMónacopascoal shelzioNo ratings yet
- Drug Development Against Tuberculosis Impact of AlkaloidsDocument41 pagesDrug Development Against Tuberculosis Impact of Alkaloidsamine ben dahsenNo ratings yet
- Proc - Bichem2006 PatersonDocument8 pagesProc - Bichem2006 PatersonRashmi RamakrishnaNo ratings yet
- Aphid Resistance in Brassica Crops - Challenges, Biotechnological Progress and Emerging PossibilitiesDocument10 pagesAphid Resistance in Brassica Crops - Challenges, Biotechnological Progress and Emerging PossibilitiesVictor Lauro Perez GarciaNo ratings yet
- 2017-R-Pina-PErez-Antimicrobial Potential of Macro and Microalgae Against Pathogenic and Spoilage Microorganisms in FoodDocument11 pages2017-R-Pina-PErez-Antimicrobial Potential of Macro and Microalgae Against Pathogenic and Spoilage Microorganisms in FoodOscar Cabrera HerraizNo ratings yet
- !antimicrobials in BeekeepingDocument11 pages!antimicrobials in BeekeepingТетяна КозицькаNo ratings yet
- DIAREDocument21 pagesDIARETutorial Kelompok 5No ratings yet
- 2019 Book AdvancesInAnimalBiotechnologyDocument556 pages2019 Book AdvancesInAnimalBiotechnologypedro hernandezNo ratings yet
- 1 s2.0 S0169409X14000246 Main PDFDocument14 pages1 s2.0 S0169409X14000246 Main PDFCesar Rodolfo Angulo DelgadoNo ratings yet
- Beil Hartz 2017Document8 pagesBeil Hartz 2017andresleonardoNo ratings yet
- Química de Los AntibióticosDocument227 pagesQuímica de Los AntibióticosArturo García FloresNo ratings yet
- Goyal 2016Document29 pagesGoyal 2016Juan Krloz CastañedaNo ratings yet
- Verma 2007Document20 pagesVerma 2007jicama SEXTO NIVELNo ratings yet
- Industrial Crops and Products: Paulo R. Ribeiro, Renato D. de Castro, Luzimar G. FernandezDocument19 pagesIndustrial Crops and Products: Paulo R. Ribeiro, Renato D. de Castro, Luzimar G. FernandezCriztIan GgomesNo ratings yet
- Chrysobalanus Icaco - A Review of Its Phytochemistry and PharmacologyDocument7 pagesChrysobalanus Icaco - A Review of Its Phytochemistry and PharmacologyPatricia Oliveira VellanoNo ratings yet
- Phitoquímica y Farmacología IcacoDocument7 pagesPhitoquímica y Farmacología IcacoJhon GiraldoNo ratings yet
- Pharmaceutically Active Secondary Metabolites of Marine ActinobacteriaDocument17 pagesPharmaceutically Active Secondary Metabolites of Marine ActinobacteriakriukkoriukNo ratings yet
- Pharmacological Potential of Selected NaDocument10 pagesPharmacological Potential of Selected NaPedro Sánchez GarcíaNo ratings yet
- Ethnopharmacological and Phytochemical Attributes of Indian Tinospora Species: A Comprehensive ReviewDocument75 pagesEthnopharmacological and Phytochemical Attributes of Indian Tinospora Species: A Comprehensive ReviewSitti Rosdiana YahyaNo ratings yet
- 3 540 33526 9Document380 pages3 540 33526 9Rituporna NathNo ratings yet
- Nichols Etal 2008Document14 pagesNichols Etal 2008Julissa O CastilloNo ratings yet
- In Vitro and in Vivo Studies of Natural Products: A Challenge For Their Valuation. The Case Study of Chamomile (Matricaria Recutita L.)Document12 pagesIn Vitro and in Vivo Studies of Natural Products: A Challenge For Their Valuation. The Case Study of Chamomile (Matricaria Recutita L.)Eduardo SantiagoNo ratings yet
- Jamu (Ginger and Kaempferia) PDFDocument23 pagesJamu (Ginger and Kaempferia) PDFuntia_sariNo ratings yet
- PAK1 Blockers Potential Therapeutics Against CO 2020 Medicine in Drug DiscoDocument5 pagesPAK1 Blockers Potential Therapeutics Against CO 2020 Medicine in Drug DiscoKhánh TườngNo ratings yet
- Chemical Constituents of Bryophytes Bio and Chemical Diversity Biological Activity and Chemosystematics 1st Edition Yoshinori AsakawaDocument54 pagesChemical Constituents of Bryophytes Bio and Chemical Diversity Biological Activity and Chemosystematics 1st Edition Yoshinori Asakawajeannie.bookman296100% (18)
- Sanjai Saxena (Auth.) - Applied Microbiology-Springer India (2015)Document201 pagesSanjai Saxena (Auth.) - Applied Microbiology-Springer India (2015)Deepika KV100% (5)
- An Evaluation of Multipurpose Oil Seed Crop For Industrial Uses (Jatropha Curcas L.) : A ReviewDocument10 pagesAn Evaluation of Multipurpose Oil Seed Crop For Industrial Uses (Jatropha Curcas L.) : A ReviewYaru HageNo ratings yet
- ESBL in WildlifeDocument93 pagesESBL in WildlifevinodhpmdNo ratings yet
- Saussurea Lappa Review 2Document12 pagesSaussurea Lappa Review 2Mulayam Singh YadavNo ratings yet
- Lactobacillos RhamnosusDocument21 pagesLactobacillos RhamnosusDoris Banila San Martin OrellanaNo ratings yet
- Beneficial Features of Plant Growth-Promoting Rhizobacteria For Improving Plant Growth and Health in Challenging Conditions: A Methodical ReviewDocument21 pagesBeneficial Features of Plant Growth-Promoting Rhizobacteria For Improving Plant Growth and Health in Challenging Conditions: A Methodical ReviewagrocursoscafNo ratings yet
- Advanced Drug Delivery Reviews: Sarah S. Tang, Anucha Apisarnthanarak, Li Yang HsuDocument11 pagesAdvanced Drug Delivery Reviews: Sarah S. Tang, Anucha Apisarnthanarak, Li Yang HsuYariahna del Carmen Rossi MaussaNo ratings yet
- 0010 1A Book Angol 01 Novenyelettan PDFDocument121 pages0010 1A Book Angol 01 Novenyelettan PDFketakeeNo ratings yet
- Micotoxinas em AlimentosDocument12 pagesMicotoxinas em AlimentosTatiany RibeiroNo ratings yet
- Bacterial Associations With LegumesDocument27 pagesBacterial Associations With LegumesJorge CorderoNo ratings yet
- Plant Growth Promoting Bacterial EndophyDocument8 pagesPlant Growth Promoting Bacterial EndophyViníciusNo ratings yet
- Amanita PhalloidesDocument15 pagesAmanita PhalloidesJair Carrillo100% (1)
- Veterinary MycologyDocument194 pagesVeterinary MycologyMuhammad Noor Bumiputra100% (1)
- Mycotoxins and Their Effects On Human and Animal HealthDocument7 pagesMycotoxins and Their Effects On Human and Animal HealthtlatilpaNo ratings yet
- 2008.guava Review PaperDocument28 pages2008.guava Review PaperRomanauli SitumorangNo ratings yet
- 1 s2.0 S016816051530194X MainDocument17 pages1 s2.0 S016816051530194X MainverticalphoebusNo ratings yet
- (SpringerBriefs in Plant Science) Münir Öztürk, Volkan Altay, Khalid Rehman Hakeem, Eren Akçiçek - Liquorice-Springer International Publishing (2017)Document150 pages(SpringerBriefs in Plant Science) Münir Öztürk, Volkan Altay, Khalid Rehman Hakeem, Eren Akçiçek - Liquorice-Springer International Publishing (2017)Rosita HandayaniNo ratings yet
- Veterinary Microbiology: Ben Adler, Alejandro de La Pen A MoctezumaDocument10 pagesVeterinary Microbiology: Ben Adler, Alejandro de La Pen A MoctezumaNaufal WildanNo ratings yet
- Adler, Pen. Leptospira and Leptospirosis. 2010 PDFDocument10 pagesAdler, Pen. Leptospira and Leptospirosis. 2010 PDFWadramys SoumsNo ratings yet
- Bernard R. Glick (Auth.) - Beneficial Plant-Bacterial Interactions-Springer International Publishing (2015)Document251 pagesBernard R. Glick (Auth.) - Beneficial Plant-Bacterial Interactions-Springer International Publishing (2015)henrique.p.guimaraesNo ratings yet
- Forensic Mycology ReviewDocument11 pagesForensic Mycology ReviewDaniel MontealegreNo ratings yet
- Factors Affecting Mushroom GrowthDocument14 pagesFactors Affecting Mushroom Growthrommel5No ratings yet
- AmfDocument165 pagesAmfJesus OcampoNo ratings yet
- Arabidopsis Thaliana Legume Lectin-Like Proteins at The InterfaceDocument139 pagesArabidopsis Thaliana Legume Lectin-Like Proteins at The Interfaceكركشندي أفندي Karkshndy afndyNo ratings yet
- 1 s2.0 S1319562X21009918 MainDocument9 pages1 s2.0 S1319562X21009918 MainmerlinakisNo ratings yet
- Bioequivalence Studies in Drug Development: Methods and ApplicationsFrom EverandBioequivalence Studies in Drug Development: Methods and ApplicationsNo ratings yet
- A Transcriptomic Survey of TheDocument20 pagesA Transcriptomic Survey of TheDwiNo ratings yet
- Action of Natural PhytosanitarDocument10 pagesAction of Natural PhytosanitarDwiNo ratings yet
- 2020-Srikaya Seed Granule Different TemperatureDocument7 pages2020-Srikaya Seed Granule Different TemperatureDwiNo ratings yet
- 2017-Justicia Adhatoda MidgutDocument11 pages2017-Justicia Adhatoda MidgutDwiNo ratings yet
- 2014-Maybe Used-Ocotea CymbarumDocument10 pages2014-Maybe Used-Ocotea CymbarumDwiNo ratings yet
- A Toxin-Binding Alkaline PhospDocument20 pagesA Toxin-Binding Alkaline PhospDwiNo ratings yet
- 2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDocument7 pages2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDwiNo ratings yet
- CHT Bfa Girls Z 0 5Document1 pageCHT Bfa Girls Z 0 5DwiNo ratings yet
- A Transformed Bacterium ExpresDocument16 pagesA Transformed Bacterium ExpresDwiNo ratings yet
- A Transgenic Approach For ContDocument8 pagesA Transgenic Approach For ContDwiNo ratings yet
- CHT Bfa Boys Z 0 5Document1 pageCHT Bfa Boys Z 0 5DwiNo ratings yet
- CHT Bfa Boys Z 2 5Document1 pageCHT Bfa Boys Z 2 5DwiNo ratings yet
- Attachment 1706 631288cdc2e9fDocument4 pagesAttachment 1706 631288cdc2e9fDwiNo ratings yet
- 2012-Eichornia Crasiper-CulexDocument7 pages2012-Eichornia Crasiper-CulexDwiNo ratings yet
- Attachment 1706 631288cdc271dDocument5 pagesAttachment 1706 631288cdc271dDwiNo ratings yet
- 2012-Fungitoxic InsecticidalDocument18 pages2012-Fungitoxic InsecticidalDwiNo ratings yet
- 2012-Garlic-Allium Sativum EndotoxinDocument11 pages2012-Garlic-Allium Sativum EndotoxinDwiNo ratings yet
- 2012-CYP450 On EarthDocument12 pages2012-CYP450 On EarthDwiNo ratings yet
- Aedes Aegypti Bacillus ThuringiensisDocument9 pagesAedes Aegypti Bacillus ThuringiensisDwiNo ratings yet
- 2012-Crimson Speckled MothDocument10 pages2012-Crimson Speckled MothDwiNo ratings yet
- 2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDocument682 pages2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDwiNo ratings yet
- 2012 Citrus Sinensis Musca DomesticaDocument8 pages2012 Citrus Sinensis Musca DomesticaDwiNo ratings yet
- 2012-Castor Bean Leaf-Spodoptera LituraDocument7 pages2012-Castor Bean Leaf-Spodoptera LituraDwiNo ratings yet
- 2012-Bti-Applied and Environmental MicrobiologyDocument6 pages2012-Bti-Applied and Environmental MicrobiologyDwiNo ratings yet
- 2012-Bti Residual TMOFDocument5 pages2012-Bti Residual TMOFDwiNo ratings yet
- 2012-Calatropis GiganteaDocument14 pages2012-Calatropis GiganteaDwiNo ratings yet
- 2012-Bti SV2, Serratia Nematodipila SV6Document7 pages2012-Bti SV2, Serratia Nematodipila SV6DwiNo ratings yet
- LESSON 1 Explain The Postulates of The Cell TheoryDocument28 pagesLESSON 1 Explain The Postulates of The Cell Theorysophia DelacruzNo ratings yet
- Science Magazine - 30 June 2023Document136 pagesScience Magazine - 30 June 2023Pablo GarcíaNo ratings yet
- Biology Notes PDF 1 Beq2woDocument373 pagesBiology Notes PDF 1 Beq2woarmanmalik5216No ratings yet
- Disorders and Diseases of Immune SystemDocument21 pagesDisorders and Diseases of Immune SystemWijesiri D WNo ratings yet
- Biomolecules WorksheetDocument4 pagesBiomolecules WorksheetJana ArafaNo ratings yet
- Mtle - Blood Banking (Revised)Document25 pagesMtle - Blood Banking (Revised)docv526No ratings yet
- How To Construct A Nature Summary Paragraph: Show" or Their Equivalent)Document1 pageHow To Construct A Nature Summary Paragraph: Show" or Their Equivalent)31 Roihan FauziNo ratings yet
- Special Staining: Intan Chairun Nisa, S.Si., M.BiotechDocument46 pagesSpecial Staining: Intan Chairun Nisa, S.Si., M.BiotechKecil Dek100% (1)
- V.O.T.E. Mock Test 1: "Champions Keep Playing Until They Get It Right."Document45 pagesV.O.T.E. Mock Test 1: "Champions Keep Playing Until They Get It Right."Miten GandhiNo ratings yet
- Genetic Disorders ProjectDocument2 pagesGenetic Disorders ProjectbarkhaNo ratings yet
- Hemifacial Microsomia: Presented by NithishDocument29 pagesHemifacial Microsomia: Presented by NithishNithish ReddyNo ratings yet
- Microbial Structures and Functions MoicroorganismsDocument390 pagesMicrobial Structures and Functions MoicroorganismssamsoatsNo ratings yet
- Impact of GMOsDocument4 pagesImpact of GMOstony montanNo ratings yet
- Dairy Cattle Breeding ManagementDocument34 pagesDairy Cattle Breeding Managementaginche amareNo ratings yet
- ANAT 2033 EoR Test 1 2015Document6 pagesANAT 2033 EoR Test 1 2015Matsiri ImmanuelNo ratings yet
- Chapter 7 Protoplast CultureDocument76 pagesChapter 7 Protoplast Culturerajiv pathakNo ratings yet
- WBC PathoDocument29 pagesWBC PathoMadhumitha ElangovanNo ratings yet
- Combining Ability Estimates in Virus Resistant and Susceptible Lines of ChilliDocument5 pagesCombining Ability Estimates in Virus Resistant and Susceptible Lines of ChilliShailendra RajanNo ratings yet
- Student Copy of GeneticsDocument122 pagesStudent Copy of GeneticsJofran TisuelaNo ratings yet
- Biology f4t2 Pp3 MsDocument4 pagesBiology f4t2 Pp3 MssamuwelNo ratings yet
- Evaluation For Module 1Document4 pagesEvaluation For Module 1Patricia PascuaNo ratings yet
- Motivational LetterDocument1 pageMotivational LetterZubair GhouriNo ratings yet
- Msczo 501Document213 pagesMsczo 501vvaishali784No ratings yet
- Tahiri (2022)Document22 pagesTahiri (2022)Jorge Andres Molina GonzalezNo ratings yet
- محاضرات ضبط الجودة أ.د. أحمد عبد المولىDocument79 pagesمحاضرات ضبط الجودة أ.د. أحمد عبد المولىMohammed AhmedNo ratings yet
- DNA Evidence in Contemporary Legal Perspective: Institutional ArticleDocument7 pagesDNA Evidence in Contemporary Legal Perspective: Institutional ArticleVijaykumar D SNo ratings yet
- 0610 BIOLOGY: MARK SCHEME For The October/November 2015 SeriesDocument14 pages0610 BIOLOGY: MARK SCHEME For The October/November 2015 SeriesKashviNo ratings yet
- Origin of Life: Earth Evolving UniverseDocument1 pageOrigin of Life: Earth Evolving UniversemateothgodNo ratings yet
- Unit 1.2 Mendelian GeneticsDocument7 pagesUnit 1.2 Mendelian GeneticsRegine Coeli Menta LansanganNo ratings yet