
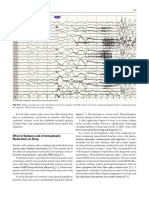
Sleep and EEG Interictal Epileptiform Abnormalities in Partial Epilepsy
Sleep and EEG Interictal Epileptiform Abnormalities in Partial Epilepsy
You might also like
- A Study in Sleep Disorders: Classification and Comprehensive AnalysisDocument18 pagesA Study in Sleep Disorders: Classification and Comprehensive AnalysisAnnapurna CMNo ratings yet
- Epilepsy Case Studies.2014 - William O. TatumDocument196 pagesEpilepsy Case Studies.2014 - William O. TatumAlexandra Mardare100% (1)
- Physiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationDocument10 pagesPhysiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationMukernas Perdossi 2021No ratings yet
- Effect of Antiepileptic Drugs On SleepDocument5 pagesEffect of Antiepileptic Drugs On SleepMukernas Perdossi 2021No ratings yet
- Relationship of Epileptic Seizures To Sleep Stage and Sleep DepthDocument6 pagesRelationship of Epileptic Seizures To Sleep Stage and Sleep Depthirsyah dwi rNo ratings yet
- Sleep & EpilepsyDocument45 pagesSleep & EpilepsyAbdul Hamid AlraiyesNo ratings yet
- Dream Recall Frequency and Content in Patients With Temporal Lobe EpilepsyDocument6 pagesDream Recall Frequency and Content in Patients With Temporal Lobe EpilepsyAnonymous zxTFUoqzklNo ratings yet
- Sleep Medicine 2015 (304 529)Document226 pagesSleep Medicine 2015 (304 529)Guillermo MoranteNo ratings yet
- Human Neurobiolog y Sleep Sleep DeprivatDocument6 pagesHuman Neurobiolog y Sleep Sleep Deprivatmelgarhoward90No ratings yet
- Sleep-Related Epilepsy: Mar Carren O, MD, PHD Santiago Ferna Ndez, MDDocument17 pagesSleep-Related Epilepsy: Mar Carren O, MD, PHD Santiago Ferna Ndez, MDMônica ScattolinNo ratings yet
- Familial Frontal Lobe Epilepsy and Its Relationship With Other Nocturnal Paroxysmal EventsDocument3 pagesFamilial Frontal Lobe Epilepsy and Its Relationship With Other Nocturnal Paroxysmal EventsxuexueNo ratings yet
- ResearchDocument10 pagesResearchkellyNo ratings yet
- All-Night EEG Power Spectral Analysis of The Cyclic Alternating Pattern at Different AgesDocument9 pagesAll-Night EEG Power Spectral Analysis of The Cyclic Alternating Pattern at Different AgespsicosmosNo ratings yet
- Journal of Sleep Research - 2012 - TERZAGHI - Dissociated Local Arousal States Underlying Essential Clinical Features ofDocument5 pagesJournal of Sleep Research - 2012 - TERZAGHI - Dissociated Local Arousal States Underlying Essential Clinical Features ofBruna PinheirosNo ratings yet
- Sleep and Epilepsy: Sejal V. Jain, MD, and Sanjeev V. Kothare, MDDocument7 pagesSleep and Epilepsy: Sejal V. Jain, MD, and Sanjeev V. Kothare, MDRezky 'kiki' Oktarianti SyahputriNo ratings yet
- Hypersomnia in An Epilepsy Patient Treated With LevetiracetamDocument2 pagesHypersomnia in An Epilepsy Patient Treated With LevetiracetamAnnisa HusainNo ratings yet
- Natural Waking and Sleep State A View From Inside Neocortical NeuronsDocument17 pagesNatural Waking and Sleep State A View From Inside Neocortical NeuronsLeslie AcevesNo ratings yet
- GeneticsDocument20 pagesGeneticsAndriNo ratings yet
- Sleep and Epilepsy Strange Bedfellows No MoreDocument26 pagesSleep and Epilepsy Strange Bedfellows No Morerifki irsyadNo ratings yet
- Journal of Sleep Research - 2016 - Schwabedal - Alpha Wave Frequency Characteristics in Health and Insomnia During SleepDocument9 pagesJournal of Sleep Research - 2016 - Schwabedal - Alpha Wave Frequency Characteristics in Health and Insomnia During SleepChisco salazar lopezNo ratings yet
- Control of Sleep and WakefulnessDocument101 pagesControl of Sleep and WakefulnessLevente BalázsNo ratings yet
- Wang 2021Document9 pagesWang 2021Aldo GuascoNo ratings yet
- 10.1097@00004691 200409000 00003Document13 pages10.1097@00004691 200409000 00003Ivan MihailovicNo ratings yet
- Peigneux Urbain Schmitz 2012 SleepandtheBrainDocument84 pagesPeigneux Urbain Schmitz 2012 SleepandtheBrainsolifugae123No ratings yet
- Borbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Document7 pagesBorbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Sebastian PereiraNo ratings yet
- C134 1945-1960 PDFDocument16 pagesC134 1945-1960 PDFstein godoy pachecoNo ratings yet
- 2003 - A Study of The Dynamic Interactions Between Sleep EEG and Heart Rate Variability in Healthy Young MenDocument10 pages2003 - A Study of The Dynamic Interactions Between Sleep EEG and Heart Rate Variability in Healthy Young MenVeronica JanethNo ratings yet
- Approaches To Staging Sleep in Polysomnographic Studies With Epileptic Activity 2003Document9 pagesApproaches To Staging Sleep in Polysomnographic Studies With Epileptic Activity 2003Bruna PinheirosNo ratings yet
- 6674 Full PDFDocument10 pages6674 Full PDFKatja GoiteNo ratings yet
- Ni Hms 815992Document18 pagesNi Hms 815992Taufik 81No ratings yet
- Parasonias em REMDocument16 pagesParasonias em REMhcrislopes5617No ratings yet
- 5-Wurts & Edgar 2000 - Circadian and Homeostatic Control of Rapid Eye Movement (REM) Sleep Promotion of REM Tendency by The Suprachiasmatic Nucleus.Document11 pages5-Wurts & Edgar 2000 - Circadian and Homeostatic Control of Rapid Eye Movement (REM) Sleep Promotion of REM Tendency by The Suprachiasmatic Nucleus.ibullerNo ratings yet
- Cohrs Et Al. (2004) - Sleep-Promoting Properties of Quetiapine in Healthy SubjectsDocument9 pagesCohrs Et Al. (2004) - Sleep-Promoting Properties of Quetiapine in Healthy SubjectsCaio RibeiroNo ratings yet
- Fpsyg 11 01662Document16 pagesFpsyg 11 01662Aldo GuascoNo ratings yet
- Sleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerriDocument9 pagesSleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerripsicosmosNo ratings yet
- Aves SuenoDocument16 pagesAves SuenoJuan Carlos Vargas HernandezNo ratings yet
- 2003 First Night Effect in Children and Adolescents Undergoing PSG For Sleep-Disordered BreathingDocument8 pages2003 First Night Effect in Children and Adolescents Undergoing PSG For Sleep-Disordered BreathingPsic ClinicaNo ratings yet
- Overview of Sleep & Sleep Disorders: Review ArticleDocument15 pagesOverview of Sleep & Sleep Disorders: Review ArticlePrima Akli SetyoNo ratings yet
- Sleep Architectural Dysfunction and Undiagnosed Obstructive SleepDocument9 pagesSleep Architectural Dysfunction and Undiagnosed Obstructive Sleepluigi.pietro.carlo.undaNo ratings yet
- Sleep and The BrainDocument5 pagesSleep and The BrainjelenaNo ratings yet
- Head Atonic Attacks. D August 21, 2012Document5 pagesHead Atonic Attacks. D August 21, 2012memoaranedaNo ratings yet
- Cote 2002Document15 pagesCote 2002Chandan ShandilNo ratings yet
- Sleep and Sleep DisordersDocument16 pagesSleep and Sleep DisordersRNo ratings yet
- Mental Activity at Sleep Onset: &. Kleitman, 19s7a, 19S7b) Has EstablishedDocument13 pagesMental Activity at Sleep Onset: &. Kleitman, 19s7a, 19S7b) Has EstablisheddoraNo ratings yet
- Epilepsia - 2018 - GibbsDocument5 pagesEpilepsia - 2018 - GibbsYenny MaharaniNo ratings yet
- Epilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CookDocument11 pagesEpilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CooknurhikmahNo ratings yet
- Jurnal HipersomniaDocument3 pagesJurnal HipersomniaKeren KaawoanNo ratings yet
- Clinical NeurophysiologyDocument5 pagesClinical Neurophysiologyleivaherre10No ratings yet
- Neurofibromin 1 Regulates Early Developmental Sleep in DrosophilaDocument9 pagesNeurofibromin 1 Regulates Early Developmental Sleep in DrosophilaFernandoNo ratings yet
- Sueño y Dolor 2019Document14 pagesSueño y Dolor 2019francescaNo ratings yet
- Zeidy 2018 Amigdala Sueño PDFDocument30 pagesZeidy 2018 Amigdala Sueño PDFFelipe JiménezNo ratings yet
- Fpsyg 11 567618Document8 pagesFpsyg 11 567618christinaceciliaaNo ratings yet
- Part II. Sleep Studies: - . - A Review of The Literature-1960-1970Document22 pagesPart II. Sleep Studies: - . - A Review of The Literature-1960-1970Melin YohanaNo ratings yet
- Sleep Disturbance and Nonmalignant Chronic PainDocument17 pagesSleep Disturbance and Nonmalignant Chronic Painveigaarthur84No ratings yet
- Neurobiology of Sleep: Madhu Kalia4Document5 pagesNeurobiology of Sleep: Madhu Kalia4Julian ReyesNo ratings yet
- Sleep Dysfunction: Rett SyndromeDocument7 pagesSleep Dysfunction: Rett SyndromeErika PratistaNo ratings yet
- Sleep Health Reciprocal Regulation of Sleep and Innate ImmunityDocument27 pagesSleep Health Reciprocal Regulation of Sleep and Innate ImmunityPolly MollyNo ratings yet
- 2003 Drug Induced Nightmares, An Etiology Based ReviewDocument9 pages2003 Drug Induced Nightmares, An Etiology Based ReviewDiego HormacheaNo ratings yet
- 1 s2.0 0031938489902436 MainDocument5 pages1 s2.0 0031938489902436 MainibullerNo ratings yet
- 7 - Deboer 2009 - Sleep and Sleep Homeostasis in Constant Darkness in The RatDocument8 pages7 - Deboer 2009 - Sleep and Sleep Homeostasis in Constant Darkness in The RatibullerNo ratings yet
- PAP VMK IndJChestDisDocument9 pagesPAP VMK IndJChestDisSmahil RNo ratings yet
- Physiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationDocument10 pagesPhysiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationMukernas Perdossi 2021No ratings yet
- Effect of Antiepileptic Drugs On SleepDocument5 pagesEffect of Antiepileptic Drugs On SleepMukernas Perdossi 2021No ratings yet
- Normal Variants Are Commonly Overread As Interictal Epileptiform AbnormalitiesDocument7 pagesNormal Variants Are Commonly Overread As Interictal Epileptiform AbnormalitiesMukernas Perdossi 2021No ratings yet
- Dyke-Davidoff-Masson-Syndrome Manifested by Seizures and Learning Difficulties: A Case ReportDocument3 pagesDyke-Davidoff-Masson-Syndrome Manifested by Seizures and Learning Difficulties: A Case ReportMukernas Perdossi 2021No ratings yet
- ICSD 3 Text Revision Supplemental MaterialDocument8 pagesICSD 3 Text Revision Supplemental Materialkepiled388No ratings yet
- Epilepsy in Children XDocument61 pagesEpilepsy in Children XSsenyonga DominicNo ratings yet
- Daftar Buku NeuroDocument4 pagesDaftar Buku NeuroDedi SutiaNo ratings yet
- TalkDocument18 pagesTalkapi-338453738No ratings yet
- PowerPoint On HeadachesDocument7 pagesPowerPoint On HeadachesMissy Wyckoff HartNo ratings yet
- UntitledDocument247 pagesUntitledalistair90No ratings yet
- Getahun Mengistu, MD, MSC, Internist, Neurologist & Specialist in Headache Medicine, Department of Neurology, AAU-MFDocument85 pagesGetahun Mengistu, MD, MSC, Internist, Neurologist & Specialist in Headache Medicine, Department of Neurology, AAU-MFashuNo ratings yet
- Anti Seizure Medications Dosing and TitrationDocument3 pagesAnti Seizure Medications Dosing and TitrationBaijuNo ratings yet
- Migraine DiaryDocument1 pageMigraine DiaryEmily KirbyNo ratings yet
- Medicamentos Essenciais para Jovens MedicosDocument2 pagesMedicamentos Essenciais para Jovens MedicosVieira MarceloNo ratings yet
- Board Ujian NeuroDocument2 pagesBoard Ujian NeuroTrisna RizkiNo ratings yet
- Vagal Nerve StimulationDocument8 pagesVagal Nerve StimulationJabraan JNo ratings yet
- Genetic Epilepsy SyndromesDocument24 pagesGenetic Epilepsy SyndromesAnali Durán CorderoNo ratings yet
- Lennox Gastaut SyndromeDocument12 pagesLennox Gastaut SyndromedulcejuvennyNo ratings yet
- Tugas 1 Neurologi Kelas 4BDocument1 pageTugas 1 Neurologi Kelas 4BWindhynrlNo ratings yet
- P&C Ep. 6. Seizure DisordersDocument36 pagesP&C Ep. 6. Seizure DisordersYahmayah von TronchinNo ratings yet
- GSAQDocument1 pageGSAQGiga HasabiNo ratings yet
- ASM Pharmacology and Clinical Efficacy 2021 PDFDocument29 pagesASM Pharmacology and Clinical Efficacy 2021 PDFIchrak GhachemNo ratings yet
- Three - Drug Combination ofDocument5 pagesThree - Drug Combination ofNaji Z. ArandiNo ratings yet
- Hassan Tonic - Clonic SeizureDocument12 pagesHassan Tonic - Clonic SeizureHassan.shehri100% (1)
- Escala D Ewport S y e Del BerlinDocument7 pagesEscala D Ewport S y e Del BerlinJuan Francisco YelaNo ratings yet
- Sharief F 2011Document14 pagesSharief F 2011wiwiNo ratings yet
- Childhood Seizure DisordersDocument57 pagesChildhood Seizure DisordersashuNo ratings yet
- EpilepsyDocument110 pagesEpilepsyAleida Ureta MorelloNo ratings yet
- Daftar Pustaka 1Document4 pagesDaftar Pustaka 1dita pangestiNo ratings yet
- Types of Seizures and Their Symptoms: SeizureDocument4 pagesTypes of Seizures and Their Symptoms: SeizureRandolph BorromeoNo ratings yet
- Epilepsy: DR Abdirahman Abib Family PhysicianDocument64 pagesEpilepsy: DR Abdirahman Abib Family PhysicianMahad IbrahemNo ratings yet
- Journal Sleep Walking 1Document7 pagesJournal Sleep Walking 1Kita SemuaNo ratings yet


























































































Download as pdf or txt
You might also like
- A Study in Sleep Disorders: Classification and Comprehensive AnalysisDocument18 pagesA Study in Sleep Disorders: Classification and Comprehensive AnalysisAnnapurna CMNo ratings yet
- Epilepsy Case Studies.2014 - William O. TatumDocument196 pagesEpilepsy Case Studies.2014 - William O. TatumAlexandra Mardare100% (1)
- Physiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationDocument10 pagesPhysiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationMukernas Perdossi 2021No ratings yet
- Effect of Antiepileptic Drugs On SleepDocument5 pagesEffect of Antiepileptic Drugs On SleepMukernas Perdossi 2021No ratings yet
- Relationship of Epileptic Seizures To Sleep Stage and Sleep DepthDocument6 pagesRelationship of Epileptic Seizures To Sleep Stage and Sleep Depthirsyah dwi rNo ratings yet
- Sleep & EpilepsyDocument45 pagesSleep & EpilepsyAbdul Hamid AlraiyesNo ratings yet
- Dream Recall Frequency and Content in Patients With Temporal Lobe EpilepsyDocument6 pagesDream Recall Frequency and Content in Patients With Temporal Lobe EpilepsyAnonymous zxTFUoqzklNo ratings yet
- Sleep Medicine 2015 (304 529)Document226 pagesSleep Medicine 2015 (304 529)Guillermo MoranteNo ratings yet
- Human Neurobiolog y Sleep Sleep DeprivatDocument6 pagesHuman Neurobiolog y Sleep Sleep Deprivatmelgarhoward90No ratings yet
- Sleep-Related Epilepsy: Mar Carren O, MD, PHD Santiago Ferna Ndez, MDDocument17 pagesSleep-Related Epilepsy: Mar Carren O, MD, PHD Santiago Ferna Ndez, MDMônica ScattolinNo ratings yet
- Familial Frontal Lobe Epilepsy and Its Relationship With Other Nocturnal Paroxysmal EventsDocument3 pagesFamilial Frontal Lobe Epilepsy and Its Relationship With Other Nocturnal Paroxysmal EventsxuexueNo ratings yet
- ResearchDocument10 pagesResearchkellyNo ratings yet
- All-Night EEG Power Spectral Analysis of The Cyclic Alternating Pattern at Different AgesDocument9 pagesAll-Night EEG Power Spectral Analysis of The Cyclic Alternating Pattern at Different AgespsicosmosNo ratings yet
- Journal of Sleep Research - 2012 - TERZAGHI - Dissociated Local Arousal States Underlying Essential Clinical Features ofDocument5 pagesJournal of Sleep Research - 2012 - TERZAGHI - Dissociated Local Arousal States Underlying Essential Clinical Features ofBruna PinheirosNo ratings yet
- Sleep and Epilepsy: Sejal V. Jain, MD, and Sanjeev V. Kothare, MDDocument7 pagesSleep and Epilepsy: Sejal V. Jain, MD, and Sanjeev V. Kothare, MDRezky 'kiki' Oktarianti SyahputriNo ratings yet
- Hypersomnia in An Epilepsy Patient Treated With LevetiracetamDocument2 pagesHypersomnia in An Epilepsy Patient Treated With LevetiracetamAnnisa HusainNo ratings yet
- Natural Waking and Sleep State A View From Inside Neocortical NeuronsDocument17 pagesNatural Waking and Sleep State A View From Inside Neocortical NeuronsLeslie AcevesNo ratings yet
- GeneticsDocument20 pagesGeneticsAndriNo ratings yet
- Sleep and Epilepsy Strange Bedfellows No MoreDocument26 pagesSleep and Epilepsy Strange Bedfellows No Morerifki irsyadNo ratings yet
- Journal of Sleep Research - 2016 - Schwabedal - Alpha Wave Frequency Characteristics in Health and Insomnia During SleepDocument9 pagesJournal of Sleep Research - 2016 - Schwabedal - Alpha Wave Frequency Characteristics in Health and Insomnia During SleepChisco salazar lopezNo ratings yet
- Control of Sleep and WakefulnessDocument101 pagesControl of Sleep and WakefulnessLevente BalázsNo ratings yet
- Wang 2021Document9 pagesWang 2021Aldo GuascoNo ratings yet
- 10.1097@00004691 200409000 00003Document13 pages10.1097@00004691 200409000 00003Ivan MihailovicNo ratings yet
- Peigneux Urbain Schmitz 2012 SleepandtheBrainDocument84 pagesPeigneux Urbain Schmitz 2012 SleepandtheBrainsolifugae123No ratings yet
- Borbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Document7 pagesBorbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Sebastian PereiraNo ratings yet
- C134 1945-1960 PDFDocument16 pagesC134 1945-1960 PDFstein godoy pachecoNo ratings yet
- 2003 - A Study of The Dynamic Interactions Between Sleep EEG and Heart Rate Variability in Healthy Young MenDocument10 pages2003 - A Study of The Dynamic Interactions Between Sleep EEG and Heart Rate Variability in Healthy Young MenVeronica JanethNo ratings yet
- Approaches To Staging Sleep in Polysomnographic Studies With Epileptic Activity 2003Document9 pagesApproaches To Staging Sleep in Polysomnographic Studies With Epileptic Activity 2003Bruna PinheirosNo ratings yet
- 6674 Full PDFDocument10 pages6674 Full PDFKatja GoiteNo ratings yet
- Ni Hms 815992Document18 pagesNi Hms 815992Taufik 81No ratings yet
- Parasonias em REMDocument16 pagesParasonias em REMhcrislopes5617No ratings yet
- 5-Wurts & Edgar 2000 - Circadian and Homeostatic Control of Rapid Eye Movement (REM) Sleep Promotion of REM Tendency by The Suprachiasmatic Nucleus.Document11 pages5-Wurts & Edgar 2000 - Circadian and Homeostatic Control of Rapid Eye Movement (REM) Sleep Promotion of REM Tendency by The Suprachiasmatic Nucleus.ibullerNo ratings yet
- Cohrs Et Al. (2004) - Sleep-Promoting Properties of Quetiapine in Healthy SubjectsDocument9 pagesCohrs Et Al. (2004) - Sleep-Promoting Properties of Quetiapine in Healthy SubjectsCaio RibeiroNo ratings yet
- Fpsyg 11 01662Document16 pagesFpsyg 11 01662Aldo GuascoNo ratings yet
- Sleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerriDocument9 pagesSleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerripsicosmosNo ratings yet
- Aves SuenoDocument16 pagesAves SuenoJuan Carlos Vargas HernandezNo ratings yet
- 2003 First Night Effect in Children and Adolescents Undergoing PSG For Sleep-Disordered BreathingDocument8 pages2003 First Night Effect in Children and Adolescents Undergoing PSG For Sleep-Disordered BreathingPsic ClinicaNo ratings yet
- Overview of Sleep & Sleep Disorders: Review ArticleDocument15 pagesOverview of Sleep & Sleep Disorders: Review ArticlePrima Akli SetyoNo ratings yet
- Sleep Architectural Dysfunction and Undiagnosed Obstructive SleepDocument9 pagesSleep Architectural Dysfunction and Undiagnosed Obstructive Sleepluigi.pietro.carlo.undaNo ratings yet
- Sleep and The BrainDocument5 pagesSleep and The BrainjelenaNo ratings yet
- Head Atonic Attacks. D August 21, 2012Document5 pagesHead Atonic Attacks. D August 21, 2012memoaranedaNo ratings yet
- Cote 2002Document15 pagesCote 2002Chandan ShandilNo ratings yet
- Sleep and Sleep DisordersDocument16 pagesSleep and Sleep DisordersRNo ratings yet
- Mental Activity at Sleep Onset: &. Kleitman, 19s7a, 19S7b) Has EstablishedDocument13 pagesMental Activity at Sleep Onset: &. Kleitman, 19s7a, 19S7b) Has EstablisheddoraNo ratings yet
- Epilepsia - 2018 - GibbsDocument5 pagesEpilepsia - 2018 - GibbsYenny MaharaniNo ratings yet
- Epilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CookDocument11 pagesEpilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CooknurhikmahNo ratings yet
- Jurnal HipersomniaDocument3 pagesJurnal HipersomniaKeren KaawoanNo ratings yet
- Clinical NeurophysiologyDocument5 pagesClinical Neurophysiologyleivaherre10No ratings yet
- Neurofibromin 1 Regulates Early Developmental Sleep in DrosophilaDocument9 pagesNeurofibromin 1 Regulates Early Developmental Sleep in DrosophilaFernandoNo ratings yet
- Sueño y Dolor 2019Document14 pagesSueño y Dolor 2019francescaNo ratings yet
- Zeidy 2018 Amigdala Sueño PDFDocument30 pagesZeidy 2018 Amigdala Sueño PDFFelipe JiménezNo ratings yet
- Fpsyg 11 567618Document8 pagesFpsyg 11 567618christinaceciliaaNo ratings yet
- Part II. Sleep Studies: - . - A Review of The Literature-1960-1970Document22 pagesPart II. Sleep Studies: - . - A Review of The Literature-1960-1970Melin YohanaNo ratings yet
- Sleep Disturbance and Nonmalignant Chronic PainDocument17 pagesSleep Disturbance and Nonmalignant Chronic Painveigaarthur84No ratings yet
- Neurobiology of Sleep: Madhu Kalia4Document5 pagesNeurobiology of Sleep: Madhu Kalia4Julian ReyesNo ratings yet
- Sleep Dysfunction: Rett SyndromeDocument7 pagesSleep Dysfunction: Rett SyndromeErika PratistaNo ratings yet
- Sleep Health Reciprocal Regulation of Sleep and Innate ImmunityDocument27 pagesSleep Health Reciprocal Regulation of Sleep and Innate ImmunityPolly MollyNo ratings yet
- 2003 Drug Induced Nightmares, An Etiology Based ReviewDocument9 pages2003 Drug Induced Nightmares, An Etiology Based ReviewDiego HormacheaNo ratings yet
- 1 s2.0 0031938489902436 MainDocument5 pages1 s2.0 0031938489902436 MainibullerNo ratings yet
- 7 - Deboer 2009 - Sleep and Sleep Homeostasis in Constant Darkness in The RatDocument8 pages7 - Deboer 2009 - Sleep and Sleep Homeostasis in Constant Darkness in The RatibullerNo ratings yet
- PAP VMK IndJChestDisDocument9 pagesPAP VMK IndJChestDisSmahil RNo ratings yet
- Physiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationDocument10 pagesPhysiological Basis: How NREM Sleep Components Can Promote and REM Sleep Components Can Suppress Seizure Discharge PropagationMukernas Perdossi 2021No ratings yet
- Effect of Antiepileptic Drugs On SleepDocument5 pagesEffect of Antiepileptic Drugs On SleepMukernas Perdossi 2021No ratings yet
- Normal Variants Are Commonly Overread As Interictal Epileptiform AbnormalitiesDocument7 pagesNormal Variants Are Commonly Overread As Interictal Epileptiform AbnormalitiesMukernas Perdossi 2021No ratings yet
- Dyke-Davidoff-Masson-Syndrome Manifested by Seizures and Learning Difficulties: A Case ReportDocument3 pagesDyke-Davidoff-Masson-Syndrome Manifested by Seizures and Learning Difficulties: A Case ReportMukernas Perdossi 2021No ratings yet
- ICSD 3 Text Revision Supplemental MaterialDocument8 pagesICSD 3 Text Revision Supplemental Materialkepiled388No ratings yet
- Epilepsy in Children XDocument61 pagesEpilepsy in Children XSsenyonga DominicNo ratings yet
- Daftar Buku NeuroDocument4 pagesDaftar Buku NeuroDedi SutiaNo ratings yet
- TalkDocument18 pagesTalkapi-338453738No ratings yet
- PowerPoint On HeadachesDocument7 pagesPowerPoint On HeadachesMissy Wyckoff HartNo ratings yet
- UntitledDocument247 pagesUntitledalistair90No ratings yet
- Getahun Mengistu, MD, MSC, Internist, Neurologist & Specialist in Headache Medicine, Department of Neurology, AAU-MFDocument85 pagesGetahun Mengistu, MD, MSC, Internist, Neurologist & Specialist in Headache Medicine, Department of Neurology, AAU-MFashuNo ratings yet
- Anti Seizure Medications Dosing and TitrationDocument3 pagesAnti Seizure Medications Dosing and TitrationBaijuNo ratings yet
- Migraine DiaryDocument1 pageMigraine DiaryEmily KirbyNo ratings yet
- Medicamentos Essenciais para Jovens MedicosDocument2 pagesMedicamentos Essenciais para Jovens MedicosVieira MarceloNo ratings yet
- Board Ujian NeuroDocument2 pagesBoard Ujian NeuroTrisna RizkiNo ratings yet
- Vagal Nerve StimulationDocument8 pagesVagal Nerve StimulationJabraan JNo ratings yet
- Genetic Epilepsy SyndromesDocument24 pagesGenetic Epilepsy SyndromesAnali Durán CorderoNo ratings yet
- Lennox Gastaut SyndromeDocument12 pagesLennox Gastaut SyndromedulcejuvennyNo ratings yet
- Tugas 1 Neurologi Kelas 4BDocument1 pageTugas 1 Neurologi Kelas 4BWindhynrlNo ratings yet
- P&C Ep. 6. Seizure DisordersDocument36 pagesP&C Ep. 6. Seizure DisordersYahmayah von TronchinNo ratings yet
- GSAQDocument1 pageGSAQGiga HasabiNo ratings yet
- ASM Pharmacology and Clinical Efficacy 2021 PDFDocument29 pagesASM Pharmacology and Clinical Efficacy 2021 PDFIchrak GhachemNo ratings yet
- Three - Drug Combination ofDocument5 pagesThree - Drug Combination ofNaji Z. ArandiNo ratings yet
- Hassan Tonic - Clonic SeizureDocument12 pagesHassan Tonic - Clonic SeizureHassan.shehri100% (1)
- Escala D Ewport S y e Del BerlinDocument7 pagesEscala D Ewport S y e Del BerlinJuan Francisco YelaNo ratings yet
- Sharief F 2011Document14 pagesSharief F 2011wiwiNo ratings yet
- Childhood Seizure DisordersDocument57 pagesChildhood Seizure DisordersashuNo ratings yet
- EpilepsyDocument110 pagesEpilepsyAleida Ureta MorelloNo ratings yet
- Daftar Pustaka 1Document4 pagesDaftar Pustaka 1dita pangestiNo ratings yet
- Types of Seizures and Their Symptoms: SeizureDocument4 pagesTypes of Seizures and Their Symptoms: SeizureRandolph BorromeoNo ratings yet
- Epilepsy: DR Abdirahman Abib Family PhysicianDocument64 pagesEpilepsy: DR Abdirahman Abib Family PhysicianMahad IbrahemNo ratings yet
- Journal Sleep Walking 1Document7 pagesJournal Sleep Walking 1Kita SemuaNo ratings yet