
Microenvironment Mapping Via Dexter Energy Transfer On Immune Cells
Microenvironment Mapping Via Dexter Energy Transfer On Immune Cells
You might also like
- The Origin and Evolutionary Biology of Pinnipeds Seals, Sea Lions, and WalrusesDocument26 pagesThe Origin and Evolutionary Biology of Pinnipeds Seals, Sea Lions, and WalrusesMaya D.No ratings yet
- Learning Activity Sheet Biotechnology 8Document11 pagesLearning Activity Sheet Biotechnology 8Jaeda BaltazarNo ratings yet
- Selectively Addressable Photogenerated Spin Qubit Pairs in DNA Hairpins (2020)Document5 pagesSelectively Addressable Photogenerated Spin Qubit Pairs in DNA Hairpins (2020)Louis Peronet VergaraNo ratings yet
- IC@N Research Projects - SPMS - May 2022Document13 pagesIC@N Research Projects - SPMS - May 2022keshavNo ratings yet
- Photosensitive Nanocarriers For Specific Delivery of Cargo Into CellsDocument12 pagesPhotosensitive Nanocarriers For Specific Delivery of Cargo Into CellsMhamed BerradaNo ratings yet
- IDE-Integrated Microneedle Arrays As Fully Biodegradable Platforms For WearableDocument4 pagesIDE-Integrated Microneedle Arrays As Fully Biodegradable Platforms For WearableNikhilNo ratings yet
- CdSe With FeSO4Document8 pagesCdSe With FeSO4Jiban MondalNo ratings yet
- Specific Detection of CA242 by Antibody-Modified Chiral Symmetric Double "N" Metasurface Biosensor - ScienceDirectDocument5 pagesSpecific Detection of CA242 by Antibody-Modified Chiral Symmetric Double "N" Metasurface Biosensor - ScienceDirectAna JuliaNo ratings yet
- Nano 2Document1 pageNano 2api-3777104No ratings yet
- Rosati, Rossi - Nonlinear Single-Particle Equations From Many-Body Lindblad (Mean Field)Document9 pagesRosati, Rossi - Nonlinear Single-Particle Equations From Many-Body Lindblad (Mean Field)Marisa UlfaNo ratings yet
- Gene Delivery Technologies With Applications in Microalgal Genetic EngineeringDocument18 pagesGene Delivery Technologies With Applications in Microalgal Genetic EngineeringmbrancovNo ratings yet
- Protein Acid NucleicDocument2 pagesProtein Acid NucleicFlorin CazanNo ratings yet
- Breaking The Barrier To Biomolecule Limit-Of-Detection Via 3D Printed Multi-Length-Scale Graphene-Coated ElectrodesDocument16 pagesBreaking The Barrier To Biomolecule Limit-Of-Detection Via 3D Printed Multi-Length-Scale Graphene-Coated ElectrodesAnahí TessaNo ratings yet
- 2024 - Broadband Thermal ImagingDocument8 pages2024 - Broadband Thermal ImaginghimansuNo ratings yet
- Dipole-Dipole Interaction in A Photonic Crystal Nanocavity: 10.1103/physreva.85.053827Document7 pagesDipole-Dipole Interaction in A Photonic Crystal Nanocavity: 10.1103/physreva.85.053827pippo50@No ratings yet
- Sensor Naqib P103412Document13 pagesSensor Naqib P103412Naqib FuadNo ratings yet
- Vietnam Journal of Chemistry - 2023 - Rooj - A Review On Characterization of Carbon Quantum DotsDocument26 pagesVietnam Journal of Chemistry - 2023 - Rooj - A Review On Characterization of Carbon Quantum DotsLCOLimaaaNo ratings yet
- MC Simulation of Radiation-Induced DNA Double Strand BreaksDocument12 pagesMC Simulation of Radiation-Induced DNA Double Strand BreaksChloe FanningNo ratings yet
- 2022 04 08 487683 FullDocument22 pages2022 04 08 487683 FullEngr. Babar AliNo ratings yet
- Bioluminescent Whole Cell Optical Ber Sensor To GenotoxicantsDocument9 pagesBioluminescent Whole Cell Optical Ber Sensor To GenotoxicantsWish NewNo ratings yet
- Wang Et Al 2007 LuminescenceDocument8 pagesWang Et Al 2007 LuminescencecarlosNo ratings yet
- Diaz Carral 2019 2D Mater. 6 045011Document9 pagesDiaz Carral 2019 2D Mater. 6 045011Ángel Díaz CarralNo ratings yet
- Mirror-Induced Re Ection in The Frequency DomainDocument9 pagesMirror-Induced Re Ection in The Frequency DomainAbelardo linoNo ratings yet
- Nonlocal Scattering Matrix Description of Anisotropic Polar HeterostructuresDocument15 pagesNonlocal Scattering Matrix Description of Anisotropic Polar Heterostructurescf fNo ratings yet
- Iturrioz-Rodríguez Et Al., 2021Document10 pagesIturrioz-Rodríguez Et Al., 2021xawadip306No ratings yet
- 2020474121, Ezeokoye Chinoso P.Document22 pages2020474121, Ezeokoye Chinoso P.ChinonyeNo ratings yet
- Engineering Luminescent Quantum Dots For in Vivo Molecular and Cellular ImagingDocument12 pagesEngineering Luminescent Quantum Dots For in Vivo Molecular and Cellular ImagingIliasa IliasNo ratings yet
- NPEE-2023-0038 Proof HiDocument36 pagesNPEE-2023-0038 Proof HiMARIA TERESA MARTINEZ MARTINEZNo ratings yet
- 33-Carbon Nanotubes TowardsDocument10 pages33-Carbon Nanotubes TowardsGABRIELNo ratings yet
- Ijms 24 00430Document8 pagesIjms 24 00430danherea7429No ratings yet
- The Wes 2007Document9 pagesThe Wes 2007Alex WongNo ratings yet
- Nanobiotechnology and BionanotechnologyDocument32 pagesNanobiotechnology and Bionanotechnologysajjad hussainNo ratings yet
- Detecting The DNA of Dengue Serotype 2 Using Aluminium Nanoparticle Doped Zinc Oxide Nanostructure: Synthesis, Analysis and CharacterizationDocument9 pagesDetecting The DNA of Dengue Serotype 2 Using Aluminium Nanoparticle Doped Zinc Oxide Nanostructure: Synthesis, Analysis and CharacterizationGayatree TripathyNo ratings yet
- (21928614 - Nanophotonics) Monolithic High-Contrast Grating Planar MicrocavitiesDocument13 pages(21928614 - Nanophotonics) Monolithic High-Contrast Grating Planar MicrocavitiesMohit SinhaNo ratings yet
- A Molecular Communication Link For Monitoring in Confined EnvironmentsDocument6 pagesA Molecular Communication Link For Monitoring in Confined EnvironmentsJohn SinhaNo ratings yet
- Chemosensors 10 00318 v2Document12 pagesChemosensors 10 00318 v2nzeyap sechieNo ratings yet
- PHD 2023-2026 Simulation of Neonicotinoid Properties in Different EnvironmentsDocument2 pagesPHD 2023-2026 Simulation of Neonicotinoid Properties in Different EnvironmentsJayanta L CNo ratings yet
- Flexible Controls of Terahertz Waves Using Coding and Programmable MetasurfacesDocument13 pagesFlexible Controls of Terahertz Waves Using Coding and Programmable MetasurfacesAnuj SharmaNo ratings yet
- Tricolor Visible Wavelength-Selective Photodegradable Hydrogel BiomaterialsDocument11 pagesTricolor Visible Wavelength-Selective Photodegradable Hydrogel Biomaterials20210901025No ratings yet
- From DNA Biosensor To Gene ChipDocument6 pagesFrom DNA Biosensor To Gene ChipramzermanNo ratings yet
- Phys Rev A 95 022337Document7 pagesPhys Rev A 95 022337Yining HuoNo ratings yet
- Direct, Electronic Microrna Detection For The Rapid Determination of Differential Expression ProfilesDocument4 pagesDirect, Electronic Microrna Detection For The Rapid Determination of Differential Expression Profilesbiosynthesis12No ratings yet
- Rapid KitDocument20 pagesRapid Kitdkp rbmNo ratings yet
- Mechanisms of Femtosecond Laser Nanosurgery of Cells and Tissues - VogelDocument33 pagesMechanisms of Femtosecond Laser Nanosurgery of Cells and Tissues - VogelBiswajit MishraNo ratings yet
- Key Parameters in Phototherapy With Gold Nanorods Using Continuous Near InfraredDocument8 pagesKey Parameters in Phototherapy With Gold Nanorods Using Continuous Near Infraredricardo23p8No ratings yet
- B.tech. Biotechnology NotesDocument14 pagesB.tech. Biotechnology NotesMudit MisraNo ratings yet
- AFM OpticalDocument9 pagesAFM OpticalNguyễn ThuNo ratings yet
- ACHOU2022Document11 pagesACHOU2022Lance ArmstrongNo ratings yet
- Sensors 21 06811 v2Document13 pagesSensors 21 06811 v2Pedro Acevedo ElgueraNo ratings yet
- Multifunctional Chitosan - Magnetic MicroneedlesDocument11 pagesMultifunctional Chitosan - Magnetic Microneedlesdavin otooleNo ratings yet
- Imaging and Characterizing Cells Using TomographyDocument28 pagesImaging and Characterizing Cells Using TomographyAJ BoonNo ratings yet
- Panda 2021Document11 pagesPanda 2021Manisha ChattopadhyayNo ratings yet
- SRRFDocument6 pagesSRRFhdgkorea19No ratings yet
- Photonic Potential of Haloarchaeal Pigment Bacteriorhodopsin For Future Electronics: A ReviewDocument9 pagesPhotonic Potential of Haloarchaeal Pigment Bacteriorhodopsin For Future Electronics: A ReviewAshwini MeiyanNo ratings yet
- Valente OC 478 126393 2021Document7 pagesValente OC 478 126393 2021paulo torresNo ratings yet
- FEBS Letters - 2020 - Turk - The Promise and The Challenges of Cryo Electron TomographyDocument19 pagesFEBS Letters - 2020 - Turk - The Promise and The Challenges of Cryo Electron Tomographyyaraloay2020No ratings yet
- The University of Lahore 1-KM Defence Road, Lahore.: Iqra Noor, Zainab Zafar, Saman RasheedDocument1 pageThe University of Lahore 1-KM Defence Road, Lahore.: Iqra Noor, Zainab Zafar, Saman RasheedKevin WilsonNo ratings yet
- Hu, 2019 PDFDocument10 pagesHu, 2019 PDFAnisa FirdausNo ratings yet
- Phototaxis Beyond Turning: Persistent Accumulation and Response Acclimation of The Microalga ChlamydomonasDocument7 pagesPhototaxis Beyond Turning: Persistent Accumulation and Response Acclimation of The Microalga Chlamydomonasalfredc20No ratings yet
- 1 s2.0 S0006349514025776 MainDocument1 page1 s2.0 S0006349514025776 MainDiego TulcanNo ratings yet
- DLL: Rina RomanoDocument6 pagesDLL: Rina RomanoRina RomanoNo ratings yet
- Topics: Nasopharyngeal CarcinomaDocument21 pagesTopics: Nasopharyngeal CarcinomaShruthi Y nursingNo ratings yet
- A Review of The CucurbitaceaeDocument15 pagesA Review of The CucurbitaceaeGuillermo BenitezNo ratings yet
- Site-Specific Recombination StrategiesDocument9 pagesSite-Specific Recombination StrategiesEdgar AbadiaNo ratings yet
- 2a. Natural Selection Simulation LabDocument3 pages2a. Natural Selection Simulation LabJames SimpsonNo ratings yet
- Complexity of Learning, Memory and Neural Processes in Evolution, Smid & Vet, 2016Document9 pagesComplexity of Learning, Memory and Neural Processes in Evolution, Smid & Vet, 2016kyudionNo ratings yet
- Cell - The Unit of Life Class 11 Notes For NEET 2024 - Free PDF DownloadDocument27 pagesCell - The Unit of Life Class 11 Notes For NEET 2024 - Free PDF Downloadathrva.sakhareNo ratings yet
- Locomotion and Movement Class 11 Notes CBSE Biology Chapter 20 (PDF)Document13 pagesLocomotion and Movement Class 11 Notes CBSE Biology Chapter 20 (PDF)ASHISH MALAVNo ratings yet
- DNA MicroarrayDocument37 pagesDNA MicroarraylordniklausNo ratings yet
- Biology Review Game - Print - QuizizzDocument5 pagesBiology Review Game - Print - QuizizzrubelliteNo ratings yet
- Transcriptome Software PaperDocument7 pagesTranscriptome Software Papershikha mittalNo ratings yet
- Peptidos AntimicrobianosDocument14 pagesPeptidos AntimicrobianosandreaNo ratings yet
- PD-L1 Expression in Triple-Negative Breast Cancer: Research ArticleDocument14 pagesPD-L1 Expression in Triple-Negative Breast Cancer: Research ArticleFebrian Parlangga MuisNo ratings yet
- UnigeneDocument7 pagesUnigeneNandni JhaNo ratings yet
- Who Classificação TO 2022 PDFDocument13 pagesWho Classificação TO 2022 PDFGrasieli RamosNo ratings yet
- Frog Embryology PDFDocument17 pagesFrog Embryology PDFHohai ConcepcionNo ratings yet
- WWW Namrata CoDocument55 pagesWWW Namrata CoMunesh SherawatNo ratings yet
- Full Download Test Bank For Genetics Analysis and Principles 4th Edition Robert Brooker PDF Full ChapterDocument34 pagesFull Download Test Bank For Genetics Analysis and Principles 4th Edition Robert Brooker PDF Full Chaptermoorpandaddyn2tmh1100% (19)
- Chemotaxonomy of FlavonoidsDocument23 pagesChemotaxonomy of Flavonoidsjaydeep raiyaniNo ratings yet
- Combined HemaDocument28 pagesCombined HemaViena Mae MaglupayNo ratings yet
- 1-259 - Ch. 3 - The CellDocument40 pages1-259 - Ch. 3 - The Celllouise navorNo ratings yet
- Genetics MCQDocument2 pagesGenetics MCQChiranjit DebbarmaNo ratings yet
- Combinatorics 2 Solutions UHSMCDocument5 pagesCombinatorics 2 Solutions UHSMCWalker KroubalkianNo ratings yet
- Activity Gene TherapyDocument3 pagesActivity Gene TherapyKent Gabriel YangaoNo ratings yet
- Most Important Topics of BiologyDocument5 pagesMost Important Topics of BiologyAshraf AliNo ratings yet
- Unit 1-Change Over Time: Daily Assignment (2/26 Wednesday)Document5 pagesUnit 1-Change Over Time: Daily Assignment (2/26 Wednesday)GnaWYrreJNo ratings yet
- Transgenic Plant and Resistance Against Abiotic StressDocument21 pagesTransgenic Plant and Resistance Against Abiotic StressMuhammad Sana UllahNo ratings yet
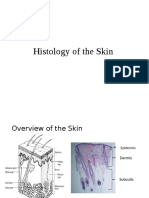
- Histology of The SkinDocument48 pagesHistology of The SkinSHARON MARIA SUNNY100% (1)

























































































Download as pdf or txt
You might also like
- The Origin and Evolutionary Biology of Pinnipeds Seals, Sea Lions, and WalrusesDocument26 pagesThe Origin and Evolutionary Biology of Pinnipeds Seals, Sea Lions, and WalrusesMaya D.No ratings yet
- Learning Activity Sheet Biotechnology 8Document11 pagesLearning Activity Sheet Biotechnology 8Jaeda BaltazarNo ratings yet
- Selectively Addressable Photogenerated Spin Qubit Pairs in DNA Hairpins (2020)Document5 pagesSelectively Addressable Photogenerated Spin Qubit Pairs in DNA Hairpins (2020)Louis Peronet VergaraNo ratings yet
- IC@N Research Projects - SPMS - May 2022Document13 pagesIC@N Research Projects - SPMS - May 2022keshavNo ratings yet
- Photosensitive Nanocarriers For Specific Delivery of Cargo Into CellsDocument12 pagesPhotosensitive Nanocarriers For Specific Delivery of Cargo Into CellsMhamed BerradaNo ratings yet
- IDE-Integrated Microneedle Arrays As Fully Biodegradable Platforms For WearableDocument4 pagesIDE-Integrated Microneedle Arrays As Fully Biodegradable Platforms For WearableNikhilNo ratings yet
- CdSe With FeSO4Document8 pagesCdSe With FeSO4Jiban MondalNo ratings yet
- Specific Detection of CA242 by Antibody-Modified Chiral Symmetric Double "N" Metasurface Biosensor - ScienceDirectDocument5 pagesSpecific Detection of CA242 by Antibody-Modified Chiral Symmetric Double "N" Metasurface Biosensor - ScienceDirectAna JuliaNo ratings yet
- Nano 2Document1 pageNano 2api-3777104No ratings yet
- Rosati, Rossi - Nonlinear Single-Particle Equations From Many-Body Lindblad (Mean Field)Document9 pagesRosati, Rossi - Nonlinear Single-Particle Equations From Many-Body Lindblad (Mean Field)Marisa UlfaNo ratings yet
- Gene Delivery Technologies With Applications in Microalgal Genetic EngineeringDocument18 pagesGene Delivery Technologies With Applications in Microalgal Genetic EngineeringmbrancovNo ratings yet
- Protein Acid NucleicDocument2 pagesProtein Acid NucleicFlorin CazanNo ratings yet
- Breaking The Barrier To Biomolecule Limit-Of-Detection Via 3D Printed Multi-Length-Scale Graphene-Coated ElectrodesDocument16 pagesBreaking The Barrier To Biomolecule Limit-Of-Detection Via 3D Printed Multi-Length-Scale Graphene-Coated ElectrodesAnahí TessaNo ratings yet
- 2024 - Broadband Thermal ImagingDocument8 pages2024 - Broadband Thermal ImaginghimansuNo ratings yet
- Dipole-Dipole Interaction in A Photonic Crystal Nanocavity: 10.1103/physreva.85.053827Document7 pagesDipole-Dipole Interaction in A Photonic Crystal Nanocavity: 10.1103/physreva.85.053827pippo50@No ratings yet
- Sensor Naqib P103412Document13 pagesSensor Naqib P103412Naqib FuadNo ratings yet
- Vietnam Journal of Chemistry - 2023 - Rooj - A Review On Characterization of Carbon Quantum DotsDocument26 pagesVietnam Journal of Chemistry - 2023 - Rooj - A Review On Characterization of Carbon Quantum DotsLCOLimaaaNo ratings yet
- MC Simulation of Radiation-Induced DNA Double Strand BreaksDocument12 pagesMC Simulation of Radiation-Induced DNA Double Strand BreaksChloe FanningNo ratings yet
- 2022 04 08 487683 FullDocument22 pages2022 04 08 487683 FullEngr. Babar AliNo ratings yet
- Bioluminescent Whole Cell Optical Ber Sensor To GenotoxicantsDocument9 pagesBioluminescent Whole Cell Optical Ber Sensor To GenotoxicantsWish NewNo ratings yet
- Wang Et Al 2007 LuminescenceDocument8 pagesWang Et Al 2007 LuminescencecarlosNo ratings yet
- Diaz Carral 2019 2D Mater. 6 045011Document9 pagesDiaz Carral 2019 2D Mater. 6 045011Ángel Díaz CarralNo ratings yet
- Mirror-Induced Re Ection in The Frequency DomainDocument9 pagesMirror-Induced Re Ection in The Frequency DomainAbelardo linoNo ratings yet
- Nonlocal Scattering Matrix Description of Anisotropic Polar HeterostructuresDocument15 pagesNonlocal Scattering Matrix Description of Anisotropic Polar Heterostructurescf fNo ratings yet
- Iturrioz-Rodríguez Et Al., 2021Document10 pagesIturrioz-Rodríguez Et Al., 2021xawadip306No ratings yet
- 2020474121, Ezeokoye Chinoso P.Document22 pages2020474121, Ezeokoye Chinoso P.ChinonyeNo ratings yet
- Engineering Luminescent Quantum Dots For in Vivo Molecular and Cellular ImagingDocument12 pagesEngineering Luminescent Quantum Dots For in Vivo Molecular and Cellular ImagingIliasa IliasNo ratings yet
- NPEE-2023-0038 Proof HiDocument36 pagesNPEE-2023-0038 Proof HiMARIA TERESA MARTINEZ MARTINEZNo ratings yet
- 33-Carbon Nanotubes TowardsDocument10 pages33-Carbon Nanotubes TowardsGABRIELNo ratings yet
- Ijms 24 00430Document8 pagesIjms 24 00430danherea7429No ratings yet
- The Wes 2007Document9 pagesThe Wes 2007Alex WongNo ratings yet
- Nanobiotechnology and BionanotechnologyDocument32 pagesNanobiotechnology and Bionanotechnologysajjad hussainNo ratings yet
- Detecting The DNA of Dengue Serotype 2 Using Aluminium Nanoparticle Doped Zinc Oxide Nanostructure: Synthesis, Analysis and CharacterizationDocument9 pagesDetecting The DNA of Dengue Serotype 2 Using Aluminium Nanoparticle Doped Zinc Oxide Nanostructure: Synthesis, Analysis and CharacterizationGayatree TripathyNo ratings yet
- (21928614 - Nanophotonics) Monolithic High-Contrast Grating Planar MicrocavitiesDocument13 pages(21928614 - Nanophotonics) Monolithic High-Contrast Grating Planar MicrocavitiesMohit SinhaNo ratings yet
- A Molecular Communication Link For Monitoring in Confined EnvironmentsDocument6 pagesA Molecular Communication Link For Monitoring in Confined EnvironmentsJohn SinhaNo ratings yet
- Chemosensors 10 00318 v2Document12 pagesChemosensors 10 00318 v2nzeyap sechieNo ratings yet
- PHD 2023-2026 Simulation of Neonicotinoid Properties in Different EnvironmentsDocument2 pagesPHD 2023-2026 Simulation of Neonicotinoid Properties in Different EnvironmentsJayanta L CNo ratings yet
- Flexible Controls of Terahertz Waves Using Coding and Programmable MetasurfacesDocument13 pagesFlexible Controls of Terahertz Waves Using Coding and Programmable MetasurfacesAnuj SharmaNo ratings yet
- Tricolor Visible Wavelength-Selective Photodegradable Hydrogel BiomaterialsDocument11 pagesTricolor Visible Wavelength-Selective Photodegradable Hydrogel Biomaterials20210901025No ratings yet
- From DNA Biosensor To Gene ChipDocument6 pagesFrom DNA Biosensor To Gene ChipramzermanNo ratings yet
- Phys Rev A 95 022337Document7 pagesPhys Rev A 95 022337Yining HuoNo ratings yet
- Direct, Electronic Microrna Detection For The Rapid Determination of Differential Expression ProfilesDocument4 pagesDirect, Electronic Microrna Detection For The Rapid Determination of Differential Expression Profilesbiosynthesis12No ratings yet
- Rapid KitDocument20 pagesRapid Kitdkp rbmNo ratings yet
- Mechanisms of Femtosecond Laser Nanosurgery of Cells and Tissues - VogelDocument33 pagesMechanisms of Femtosecond Laser Nanosurgery of Cells and Tissues - VogelBiswajit MishraNo ratings yet
- Key Parameters in Phototherapy With Gold Nanorods Using Continuous Near InfraredDocument8 pagesKey Parameters in Phototherapy With Gold Nanorods Using Continuous Near Infraredricardo23p8No ratings yet
- B.tech. Biotechnology NotesDocument14 pagesB.tech. Biotechnology NotesMudit MisraNo ratings yet
- AFM OpticalDocument9 pagesAFM OpticalNguyễn ThuNo ratings yet
- ACHOU2022Document11 pagesACHOU2022Lance ArmstrongNo ratings yet
- Sensors 21 06811 v2Document13 pagesSensors 21 06811 v2Pedro Acevedo ElgueraNo ratings yet
- Multifunctional Chitosan - Magnetic MicroneedlesDocument11 pagesMultifunctional Chitosan - Magnetic Microneedlesdavin otooleNo ratings yet
- Imaging and Characterizing Cells Using TomographyDocument28 pagesImaging and Characterizing Cells Using TomographyAJ BoonNo ratings yet
- Panda 2021Document11 pagesPanda 2021Manisha ChattopadhyayNo ratings yet
- SRRFDocument6 pagesSRRFhdgkorea19No ratings yet
- Photonic Potential of Haloarchaeal Pigment Bacteriorhodopsin For Future Electronics: A ReviewDocument9 pagesPhotonic Potential of Haloarchaeal Pigment Bacteriorhodopsin For Future Electronics: A ReviewAshwini MeiyanNo ratings yet
- Valente OC 478 126393 2021Document7 pagesValente OC 478 126393 2021paulo torresNo ratings yet
- FEBS Letters - 2020 - Turk - The Promise and The Challenges of Cryo Electron TomographyDocument19 pagesFEBS Letters - 2020 - Turk - The Promise and The Challenges of Cryo Electron Tomographyyaraloay2020No ratings yet
- The University of Lahore 1-KM Defence Road, Lahore.: Iqra Noor, Zainab Zafar, Saman RasheedDocument1 pageThe University of Lahore 1-KM Defence Road, Lahore.: Iqra Noor, Zainab Zafar, Saman RasheedKevin WilsonNo ratings yet
- Hu, 2019 PDFDocument10 pagesHu, 2019 PDFAnisa FirdausNo ratings yet
- Phototaxis Beyond Turning: Persistent Accumulation and Response Acclimation of The Microalga ChlamydomonasDocument7 pagesPhototaxis Beyond Turning: Persistent Accumulation and Response Acclimation of The Microalga Chlamydomonasalfredc20No ratings yet
- 1 s2.0 S0006349514025776 MainDocument1 page1 s2.0 S0006349514025776 MainDiego TulcanNo ratings yet
- DLL: Rina RomanoDocument6 pagesDLL: Rina RomanoRina RomanoNo ratings yet
- Topics: Nasopharyngeal CarcinomaDocument21 pagesTopics: Nasopharyngeal CarcinomaShruthi Y nursingNo ratings yet
- A Review of The CucurbitaceaeDocument15 pagesA Review of The CucurbitaceaeGuillermo BenitezNo ratings yet
- Site-Specific Recombination StrategiesDocument9 pagesSite-Specific Recombination StrategiesEdgar AbadiaNo ratings yet
- 2a. Natural Selection Simulation LabDocument3 pages2a. Natural Selection Simulation LabJames SimpsonNo ratings yet
- Complexity of Learning, Memory and Neural Processes in Evolution, Smid & Vet, 2016Document9 pagesComplexity of Learning, Memory and Neural Processes in Evolution, Smid & Vet, 2016kyudionNo ratings yet
- Cell - The Unit of Life Class 11 Notes For NEET 2024 - Free PDF DownloadDocument27 pagesCell - The Unit of Life Class 11 Notes For NEET 2024 - Free PDF Downloadathrva.sakhareNo ratings yet
- Locomotion and Movement Class 11 Notes CBSE Biology Chapter 20 (PDF)Document13 pagesLocomotion and Movement Class 11 Notes CBSE Biology Chapter 20 (PDF)ASHISH MALAVNo ratings yet
- DNA MicroarrayDocument37 pagesDNA MicroarraylordniklausNo ratings yet
- Biology Review Game - Print - QuizizzDocument5 pagesBiology Review Game - Print - QuizizzrubelliteNo ratings yet
- Transcriptome Software PaperDocument7 pagesTranscriptome Software Papershikha mittalNo ratings yet
- Peptidos AntimicrobianosDocument14 pagesPeptidos AntimicrobianosandreaNo ratings yet
- PD-L1 Expression in Triple-Negative Breast Cancer: Research ArticleDocument14 pagesPD-L1 Expression in Triple-Negative Breast Cancer: Research ArticleFebrian Parlangga MuisNo ratings yet
- UnigeneDocument7 pagesUnigeneNandni JhaNo ratings yet
- Who Classificação TO 2022 PDFDocument13 pagesWho Classificação TO 2022 PDFGrasieli RamosNo ratings yet
- Frog Embryology PDFDocument17 pagesFrog Embryology PDFHohai ConcepcionNo ratings yet
- WWW Namrata CoDocument55 pagesWWW Namrata CoMunesh SherawatNo ratings yet
- Full Download Test Bank For Genetics Analysis and Principles 4th Edition Robert Brooker PDF Full ChapterDocument34 pagesFull Download Test Bank For Genetics Analysis and Principles 4th Edition Robert Brooker PDF Full Chaptermoorpandaddyn2tmh1100% (19)
- Chemotaxonomy of FlavonoidsDocument23 pagesChemotaxonomy of Flavonoidsjaydeep raiyaniNo ratings yet
- Combined HemaDocument28 pagesCombined HemaViena Mae MaglupayNo ratings yet
- 1-259 - Ch. 3 - The CellDocument40 pages1-259 - Ch. 3 - The Celllouise navorNo ratings yet
- Genetics MCQDocument2 pagesGenetics MCQChiranjit DebbarmaNo ratings yet
- Combinatorics 2 Solutions UHSMCDocument5 pagesCombinatorics 2 Solutions UHSMCWalker KroubalkianNo ratings yet
- Activity Gene TherapyDocument3 pagesActivity Gene TherapyKent Gabriel YangaoNo ratings yet
- Most Important Topics of BiologyDocument5 pagesMost Important Topics of BiologyAshraf AliNo ratings yet
- Unit 1-Change Over Time: Daily Assignment (2/26 Wednesday)Document5 pagesUnit 1-Change Over Time: Daily Assignment (2/26 Wednesday)GnaWYrreJNo ratings yet
- Transgenic Plant and Resistance Against Abiotic StressDocument21 pagesTransgenic Plant and Resistance Against Abiotic StressMuhammad Sana UllahNo ratings yet
- Histology of The SkinDocument48 pagesHistology of The SkinSHARON MARIA SUNNY100% (1)