
Veterinary Immunology and Immunopathology: A. Beineke, C. Puff, F. Seehusen, W. Baumga Rtner
Veterinary Immunology and Immunopathology: A. Beineke, C. Puff, F. Seehusen, W. Baumga Rtner
You might also like
- Immunology - Website QuestionsDocument100 pagesImmunology - Website QuestionsMohammed AlMujaini75% (12)
- Pathophysiology Exam QuestionsDocument11 pagesPathophysiology Exam Questionsjimmy100% (2)
- Immunology Uworld Notes (Step 1)Document12 pagesImmunology Uworld Notes (Step 1)Burkitt's LymphomaNo ratings yet
- VII MostDown 1Document28 pagesVII MostDown 1Orange LemonNo ratings yet
- Epidemiology of Infections in Cancer Patients: Teresa R. ZembowerDocument48 pagesEpidemiology of Infections in Cancer Patients: Teresa R. ZembowerArben TecsonNo ratings yet
- Chachage Mkunde PDFDocument157 pagesChachage Mkunde PDFMegbaruNo ratings yet
- Textbook Malaria Immune Response To Infection and Vaccination 1St Edition Maria M Mota Ebook All Chapter PDFDocument53 pagesTextbook Malaria Immune Response To Infection and Vaccination 1St Edition Maria M Mota Ebook All Chapter PDFjoyce.cummins304100% (12)
- Immunity To Extracellular and Intracellular Bacteria 2 3 2Document8 pagesImmunity To Extracellular and Intracellular Bacteria 2 3 2Habtamu Temesgen SdaNo ratings yet
- 2 Pathophysiology of HIVDocument8 pages2 Pathophysiology of HIVRaga ManduaruNo ratings yet
- Poliovirus, Pathogenesis of Poliomyelitis, and Apoptosis: Current Topics in Microbiology and Immunology February 2005Document33 pagesPoliovirus, Pathogenesis of Poliomyelitis, and Apoptosis: Current Topics in Microbiology and Immunology February 2005Ngurah BodhinathaNo ratings yet
- 2 Pa Tho Physiology of HIVDocument8 pages2 Pa Tho Physiology of HIVLeepin TanNo ratings yet
- Non-inflammatory immunology: An introduction to the immune system and its pathologiesFrom EverandNon-inflammatory immunology: An introduction to the immune system and its pathologiesNo ratings yet
- Immunology TuteDocument9 pagesImmunology TuteNimesha GammanpilaNo ratings yet
- Natural Immunomodulators Treat Cytokine Storm in Covid, Sepsis, Etc.Document17 pagesNatural Immunomodulators Treat Cytokine Storm in Covid, Sepsis, Etc.Sherry PhippsNo ratings yet
- Immunity To Microbes - IDocument23 pagesImmunity To Microbes - Ila.instagramawaNo ratings yet
- Autoimmune Diseases of Oral CavityDocument52 pagesAutoimmune Diseases of Oral Cavitylakshmi k sNo ratings yet
- Historical Insights Into CytokinesDocument13 pagesHistorical Insights Into CytokinesRickySaptarshi SahaNo ratings yet
- Carlberg C. Molecular Immunology. How Science Works 2022Document228 pagesCarlberg C. Molecular Immunology. How Science Works 2022Philip Rodríguez GarcíaNo ratings yet
- Pasteurella Multocida and Immune CellsDocument20 pagesPasteurella Multocida and Immune CellsOmar Ramirez JesúsNo ratings yet
- Nelson 2017Document28 pagesNelson 2017Santiago GonzalezNo ratings yet
- Human Health and DiseaseDocument12 pagesHuman Health and DiseasePratham PatelNo ratings yet
- Of Sepsis: New Concepts and Implications For Science, Medicine, and The Future: PathogenesisDocument6 pagesOf Sepsis: New Concepts and Implications For Science, Medicine, and The Future: PathogenesisAndyan Adlu PrasetyajiNo ratings yet
- Humoral Immune Mechanisms Involved in Protective and Pathological MARKEDDocument14 pagesHumoral Immune Mechanisms Involved in Protective and Pathological MARKEDLayla LopesNo ratings yet
- Irwansyah Nur Oktafian - 180342618025 - Immunology TasksDocument5 pagesIrwansyah Nur Oktafian - 180342618025 - Immunology TasksIRWANSYAH NURNo ratings yet
- Basic ImmunologyDocument29 pagesBasic ImmunologyHema Duddukuri0% (1)
- +2 Bio Zoo em 7 12 LessonsDocument30 pages+2 Bio Zoo em 7 12 LessonsInnovative IndustryNo ratings yet
- Kornman, Page, Tonetti 1997-Periodontology - 2000Document22 pagesKornman, Page, Tonetti 1997-Periodontology - 2000Liliana Miranda ANo ratings yet
- Morales J. The Innate Immune System in Health... Vol 1. 2022Document328 pagesMorales J. The Innate Immune System in Health... Vol 1. 2022Philip Rodríguez GarcíaNo ratings yet
- Immunological Pathogenesis and Treatment of SystemDocument12 pagesImmunological Pathogenesis and Treatment of SystemlousianaNo ratings yet
- +2 Bio-Zoo-Em 7-12Document30 pages+2 Bio-Zoo-Em 7-12aj techNo ratings yet
- Ojsadmin, 1491Document4 pagesOjsadmin, 1491Cristian PeñaNo ratings yet
- Helminth Infections - Recognition and Modulation of The Immune Response by Innate Immune CellsDocument12 pagesHelminth Infections - Recognition and Modulation of The Immune Response by Innate Immune CellsponbohacopNo ratings yet
- The Immune System: Back To The BasicsDocument37 pagesThe Immune System: Back To The BasicsHani GowaiNo ratings yet
- Cytoquine Storm and SepsisDocument12 pagesCytoquine Storm and SepsisEduardo ChanonaNo ratings yet
- Psoriasis Pathophysiology Current ConceptsofpathogenesisDocument7 pagesPsoriasis Pathophysiology Current ConceptsofpathogenesisAnnaNo ratings yet
- Diseases of The Immune SystemDocument65 pagesDiseases of The Immune Systemanon_62816775No ratings yet
- 1 s2.0 S1569199317308755 MainDocument10 pages1 s2.0 S1569199317308755 MainraniahNo ratings yet
- Synergistic Effects of Ageing and Stress On Neutrophil FunctionDocument21 pagesSynergistic Effects of Ageing and Stress On Neutrophil Functionbenefits35No ratings yet
- Mascelli NoDocument10 pagesMascelli NoAung SoeNo ratings yet
- Inmunopatogenesis Del LupusDocument13 pagesInmunopatogenesis Del LupusPdfzNo ratings yet
- Immunology of Periodontal DiseasesDocument9 pagesImmunology of Periodontal DiseasesoladunniNo ratings yet
- Low CD4 in HIVDocument27 pagesLow CD4 in HIVwinwinwinaNo ratings yet
- Virology NotesDocument81 pagesVirology NotesraulNo ratings yet
- Immunosenescence, Inflammaging and Cell SenescenceDocument63 pagesImmunosenescence, Inflammaging and Cell SenescenceArthur YanezNo ratings yet
- Immune System To Agent InfectionDocument70 pagesImmune System To Agent Infectionliza yunitaNo ratings yet
- Immunological Pathogenesis and Treatment of Systemic Lupus ErythematosusDocument12 pagesImmunological Pathogenesis and Treatment of Systemic Lupus ErythematosusLizeth Rincon DelgadoNo ratings yet
- Rajiv Gandhi Institute of Veterinary Education and Research Kurumbapet, Puducherry - 605 007Document11 pagesRajiv Gandhi Institute of Veterinary Education and Research Kurumbapet, Puducherry - 605 007Kavi BharathyNo ratings yet
- Viral Structure NotesDocument9 pagesViral Structure Notesantonia putriNo ratings yet
- Immunological Aspects IAL CH 7Document79 pagesImmunological Aspects IAL CH 7SheilaNo ratings yet
- vt59.2708 21406740150 - 885398426264138 - 2308855381410239775 - n.pdf4. Infectious Inflammatory and ImmuDocument180 pagesvt59.2708 21406740150 - 885398426264138 - 2308855381410239775 - n.pdf4. Infectious Inflammatory and ImmuswsnkkcgbhNo ratings yet
- 2012 Innate Immunosenescence Effect of Aging On Cells and Receptors of The Innate Immune System in Humans. Seminars in ImmunologyDocument11 pages2012 Innate Immunosenescence Effect of Aging On Cells and Receptors of The Innate Immune System in Humans. Seminars in ImmunologyManu TrejosNo ratings yet
- Immunological Responses To Bacteria in The Human BodyDocument13 pagesImmunological Responses To Bacteria in The Human BodyDivine AffluenceNo ratings yet
- I Mmuni LogyDocument12 pagesI Mmuni Logymessebo90No ratings yet
- Rheumatoid Arthritis: Recent Advances On Its Etiology, Role of Cytokines and PharmacotherapyDocument19 pagesRheumatoid Arthritis: Recent Advances On Its Etiology, Role of Cytokines and PharmacotherapyRachel Lalaine Marie SialanaNo ratings yet
- Inflammation CCDocument72 pagesInflammation CCmulatumeleseNo ratings yet
- Mouse Models of Inflammatory Bowel Disease: Stefan Wirtz, Markus F. NeurathDocument11 pagesMouse Models of Inflammatory Bowel Disease: Stefan Wirtz, Markus F. NeurathChristianDuarteVarelaNo ratings yet
- Ebook Cellular and Molecular Immunology 8Th Edition Abbas Test Bank Full Chapter PDFDocument26 pagesEbook Cellular and Molecular Immunology 8Th Edition Abbas Test Bank Full Chapter PDFDavidLeekemt100% (13)
- RegulDocument3 pagesRegulBestiaNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- The Microbiome in Rheumatic Diseases and InfectionFrom EverandThe Microbiome in Rheumatic Diseases and InfectionGaafar RagabNo ratings yet
- HPV-Infection and Cancers of the Skin - Diagnostics and TherapyFrom EverandHPV-Infection and Cancers of the Skin - Diagnostics and TherapyRating: 5 out of 5 stars5/5 (1)
- Lymphatic Drainage of Head & NeckDocument96 pagesLymphatic Drainage of Head & NeckRoopa PatilNo ratings yet
- Treating Breast Cancer: Local TreatmentsDocument99 pagesTreating Breast Cancer: Local Treatmentssuci triana putriNo ratings yet
- Anatomy and Physiology of Lymphatic SystemDocument24 pagesAnatomy and Physiology of Lymphatic Systemamber tariqNo ratings yet
- LYMPHOMADocument12 pagesLYMPHOMAश्रीकृष्ण हेङ्गजूNo ratings yet
- Lymphatic SystemDocument22 pagesLymphatic SystemdrynwhylNo ratings yet
- Examination of Lymph Nodes of Head and NeckDocument58 pagesExamination of Lymph Nodes of Head and NeckabdulrazaqNo ratings yet
- Micronatomy pt2Document35 pagesMicronatomy pt2poopziNo ratings yet
- PHC411 - Practical 3 - Oct 2021 - StudentsDocument14 pagesPHC411 - Practical 3 - Oct 2021 - StudentsALISYA SOPHIA MOHAMMAD ABU SHAHID CHRISNo ratings yet
- Advanced Piezoelectric Materials Science and Technology 2nd Edition Kenji Uchino Full ChapterDocument24 pagesAdvanced Piezoelectric Materials Science and Technology 2nd Edition Kenji Uchino Full Chapterwejzagough100% (3)
- 24 Lecture PresentationDocument85 pages24 Lecture PresentationWide PutinNo ratings yet
- Abreastcancerstory Breastcancertreatmentreview 100624205948 Phpapp01Document552 pagesAbreastcancerstory Breastcancertreatmentreview 100624205948 Phpapp01muhammad yamin100% (1)
- The SPLENOMEGALY Power Point Dr. CosmescuDocument29 pagesThe SPLENOMEGALY Power Point Dr. CosmescuNisha PillaiNo ratings yet
- Crystal Shipping, Inc. vs. Natividad, 473 SCRA 559 (2005Document15 pagesCrystal Shipping, Inc. vs. Natividad, 473 SCRA 559 (2005Mei SuNo ratings yet
- HistologyDocument18 pagesHistologyGlavinorNo ratings yet
- Immunology Tonsil 2Document31 pagesImmunology Tonsil 2ariienndrrahanniie100% (1)
- Leadership Initiative To Enrich The Hiv/aids Education For Senior High SchoolsDocument21 pagesLeadership Initiative To Enrich The Hiv/aids Education For Senior High SchoolsIshika JainNo ratings yet
- Ultrasound of Thyroid, Parathyroid Glands and Neck Lymph Nodes - SolbiatiDocument14 pagesUltrasound of Thyroid, Parathyroid Glands and Neck Lymph Nodes - Solbiatidrmoscalin8774No ratings yet
- Immune System Overview - Understanding The Immune SystemDocument60 pagesImmune System Overview - Understanding The Immune Systemlourdesbr100% (2)
- LimfadenitisDocument24 pagesLimfadenitisrahmah ningsihNo ratings yet
- Лекция 5Document27 pagesЛекция 5Oriyomi OdekunleNo ratings yet
- 4 2Document3 pages4 2Stefia AisyahNo ratings yet
- LyphomaDocument7 pagesLyphomaNambuye Midyero AhmedNo ratings yet
- 1 Крок 1 ЛС KROK 2007 2017 English BiochemistryDocument480 pages1 Крок 1 ЛС KROK 2007 2017 English BiochemistryAbhani MøhitNo ratings yet
- How I Evaluate The Neck Lymph NodesDocument29 pagesHow I Evaluate The Neck Lymph NodesMartín Avellaneda SantoméNo ratings yet
- 12th Zoology Practical Study Material English MediumDocument24 pages12th Zoology Practical Study Material English MediumKarthika KarthikaNo ratings yet
- Microscopic Descriptive TechniquesDocument13 pagesMicroscopic Descriptive TechniquesJill PNo ratings yet
- Clinical Feature of Lymphnode EnlargmentDocument47 pagesClinical Feature of Lymphnode Enlargmentjenny girlNo ratings yet
- Non HodgkinsDocument2 pagesNon HodgkinstapNo ratings yet
- Laboratory Methodsin ImmnunologyDocument58 pagesLaboratory Methodsin Immnunologyadi pNo ratings yet
































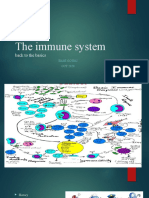









































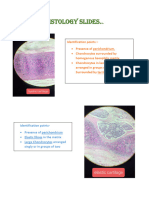














Download as pdf or txt
You might also like
- Immunology - Website QuestionsDocument100 pagesImmunology - Website QuestionsMohammed AlMujaini75% (12)
- Pathophysiology Exam QuestionsDocument11 pagesPathophysiology Exam Questionsjimmy100% (2)
- Immunology Uworld Notes (Step 1)Document12 pagesImmunology Uworld Notes (Step 1)Burkitt's LymphomaNo ratings yet
- VII MostDown 1Document28 pagesVII MostDown 1Orange LemonNo ratings yet
- Epidemiology of Infections in Cancer Patients: Teresa R. ZembowerDocument48 pagesEpidemiology of Infections in Cancer Patients: Teresa R. ZembowerArben TecsonNo ratings yet
- Chachage Mkunde PDFDocument157 pagesChachage Mkunde PDFMegbaruNo ratings yet
- Textbook Malaria Immune Response To Infection and Vaccination 1St Edition Maria M Mota Ebook All Chapter PDFDocument53 pagesTextbook Malaria Immune Response To Infection and Vaccination 1St Edition Maria M Mota Ebook All Chapter PDFjoyce.cummins304100% (12)
- Immunity To Extracellular and Intracellular Bacteria 2 3 2Document8 pagesImmunity To Extracellular and Intracellular Bacteria 2 3 2Habtamu Temesgen SdaNo ratings yet
- 2 Pathophysiology of HIVDocument8 pages2 Pathophysiology of HIVRaga ManduaruNo ratings yet
- Poliovirus, Pathogenesis of Poliomyelitis, and Apoptosis: Current Topics in Microbiology and Immunology February 2005Document33 pagesPoliovirus, Pathogenesis of Poliomyelitis, and Apoptosis: Current Topics in Microbiology and Immunology February 2005Ngurah BodhinathaNo ratings yet
- 2 Pa Tho Physiology of HIVDocument8 pages2 Pa Tho Physiology of HIVLeepin TanNo ratings yet
- Non-inflammatory immunology: An introduction to the immune system and its pathologiesFrom EverandNon-inflammatory immunology: An introduction to the immune system and its pathologiesNo ratings yet
- Immunology TuteDocument9 pagesImmunology TuteNimesha GammanpilaNo ratings yet
- Natural Immunomodulators Treat Cytokine Storm in Covid, Sepsis, Etc.Document17 pagesNatural Immunomodulators Treat Cytokine Storm in Covid, Sepsis, Etc.Sherry PhippsNo ratings yet
- Immunity To Microbes - IDocument23 pagesImmunity To Microbes - Ila.instagramawaNo ratings yet
- Autoimmune Diseases of Oral CavityDocument52 pagesAutoimmune Diseases of Oral Cavitylakshmi k sNo ratings yet
- Historical Insights Into CytokinesDocument13 pagesHistorical Insights Into CytokinesRickySaptarshi SahaNo ratings yet
- Carlberg C. Molecular Immunology. How Science Works 2022Document228 pagesCarlberg C. Molecular Immunology. How Science Works 2022Philip Rodríguez GarcíaNo ratings yet
- Pasteurella Multocida and Immune CellsDocument20 pagesPasteurella Multocida and Immune CellsOmar Ramirez JesúsNo ratings yet
- Nelson 2017Document28 pagesNelson 2017Santiago GonzalezNo ratings yet
- Human Health and DiseaseDocument12 pagesHuman Health and DiseasePratham PatelNo ratings yet
- Of Sepsis: New Concepts and Implications For Science, Medicine, and The Future: PathogenesisDocument6 pagesOf Sepsis: New Concepts and Implications For Science, Medicine, and The Future: PathogenesisAndyan Adlu PrasetyajiNo ratings yet
- Humoral Immune Mechanisms Involved in Protective and Pathological MARKEDDocument14 pagesHumoral Immune Mechanisms Involved in Protective and Pathological MARKEDLayla LopesNo ratings yet
- Irwansyah Nur Oktafian - 180342618025 - Immunology TasksDocument5 pagesIrwansyah Nur Oktafian - 180342618025 - Immunology TasksIRWANSYAH NURNo ratings yet
- Basic ImmunologyDocument29 pagesBasic ImmunologyHema Duddukuri0% (1)
- +2 Bio Zoo em 7 12 LessonsDocument30 pages+2 Bio Zoo em 7 12 LessonsInnovative IndustryNo ratings yet
- Kornman, Page, Tonetti 1997-Periodontology - 2000Document22 pagesKornman, Page, Tonetti 1997-Periodontology - 2000Liliana Miranda ANo ratings yet
- Morales J. The Innate Immune System in Health... Vol 1. 2022Document328 pagesMorales J. The Innate Immune System in Health... Vol 1. 2022Philip Rodríguez GarcíaNo ratings yet
- Immunological Pathogenesis and Treatment of SystemDocument12 pagesImmunological Pathogenesis and Treatment of SystemlousianaNo ratings yet
- +2 Bio-Zoo-Em 7-12Document30 pages+2 Bio-Zoo-Em 7-12aj techNo ratings yet
- Ojsadmin, 1491Document4 pagesOjsadmin, 1491Cristian PeñaNo ratings yet
- Helminth Infections - Recognition and Modulation of The Immune Response by Innate Immune CellsDocument12 pagesHelminth Infections - Recognition and Modulation of The Immune Response by Innate Immune CellsponbohacopNo ratings yet
- The Immune System: Back To The BasicsDocument37 pagesThe Immune System: Back To The BasicsHani GowaiNo ratings yet
- Cytoquine Storm and SepsisDocument12 pagesCytoquine Storm and SepsisEduardo ChanonaNo ratings yet
- Psoriasis Pathophysiology Current ConceptsofpathogenesisDocument7 pagesPsoriasis Pathophysiology Current ConceptsofpathogenesisAnnaNo ratings yet
- Diseases of The Immune SystemDocument65 pagesDiseases of The Immune Systemanon_62816775No ratings yet
- 1 s2.0 S1569199317308755 MainDocument10 pages1 s2.0 S1569199317308755 MainraniahNo ratings yet
- Synergistic Effects of Ageing and Stress On Neutrophil FunctionDocument21 pagesSynergistic Effects of Ageing and Stress On Neutrophil Functionbenefits35No ratings yet
- Mascelli NoDocument10 pagesMascelli NoAung SoeNo ratings yet
- Inmunopatogenesis Del LupusDocument13 pagesInmunopatogenesis Del LupusPdfzNo ratings yet
- Immunology of Periodontal DiseasesDocument9 pagesImmunology of Periodontal DiseasesoladunniNo ratings yet
- Low CD4 in HIVDocument27 pagesLow CD4 in HIVwinwinwinaNo ratings yet
- Virology NotesDocument81 pagesVirology NotesraulNo ratings yet
- Immunosenescence, Inflammaging and Cell SenescenceDocument63 pagesImmunosenescence, Inflammaging and Cell SenescenceArthur YanezNo ratings yet
- Immune System To Agent InfectionDocument70 pagesImmune System To Agent Infectionliza yunitaNo ratings yet
- Immunological Pathogenesis and Treatment of Systemic Lupus ErythematosusDocument12 pagesImmunological Pathogenesis and Treatment of Systemic Lupus ErythematosusLizeth Rincon DelgadoNo ratings yet
- Rajiv Gandhi Institute of Veterinary Education and Research Kurumbapet, Puducherry - 605 007Document11 pagesRajiv Gandhi Institute of Veterinary Education and Research Kurumbapet, Puducherry - 605 007Kavi BharathyNo ratings yet
- Viral Structure NotesDocument9 pagesViral Structure Notesantonia putriNo ratings yet
- Immunological Aspects IAL CH 7Document79 pagesImmunological Aspects IAL CH 7SheilaNo ratings yet
- vt59.2708 21406740150 - 885398426264138 - 2308855381410239775 - n.pdf4. Infectious Inflammatory and ImmuDocument180 pagesvt59.2708 21406740150 - 885398426264138 - 2308855381410239775 - n.pdf4. Infectious Inflammatory and ImmuswsnkkcgbhNo ratings yet
- 2012 Innate Immunosenescence Effect of Aging On Cells and Receptors of The Innate Immune System in Humans. Seminars in ImmunologyDocument11 pages2012 Innate Immunosenescence Effect of Aging On Cells and Receptors of The Innate Immune System in Humans. Seminars in ImmunologyManu TrejosNo ratings yet
- Immunological Responses To Bacteria in The Human BodyDocument13 pagesImmunological Responses To Bacteria in The Human BodyDivine AffluenceNo ratings yet
- I Mmuni LogyDocument12 pagesI Mmuni Logymessebo90No ratings yet
- Rheumatoid Arthritis: Recent Advances On Its Etiology, Role of Cytokines and PharmacotherapyDocument19 pagesRheumatoid Arthritis: Recent Advances On Its Etiology, Role of Cytokines and PharmacotherapyRachel Lalaine Marie SialanaNo ratings yet
- Inflammation CCDocument72 pagesInflammation CCmulatumeleseNo ratings yet
- Mouse Models of Inflammatory Bowel Disease: Stefan Wirtz, Markus F. NeurathDocument11 pagesMouse Models of Inflammatory Bowel Disease: Stefan Wirtz, Markus F. NeurathChristianDuarteVarelaNo ratings yet
- Ebook Cellular and Molecular Immunology 8Th Edition Abbas Test Bank Full Chapter PDFDocument26 pagesEbook Cellular and Molecular Immunology 8Th Edition Abbas Test Bank Full Chapter PDFDavidLeekemt100% (13)
- RegulDocument3 pagesRegulBestiaNo ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- The Microbiome in Rheumatic Diseases and InfectionFrom EverandThe Microbiome in Rheumatic Diseases and InfectionGaafar RagabNo ratings yet
- HPV-Infection and Cancers of the Skin - Diagnostics and TherapyFrom EverandHPV-Infection and Cancers of the Skin - Diagnostics and TherapyRating: 5 out of 5 stars5/5 (1)
- Lymphatic Drainage of Head & NeckDocument96 pagesLymphatic Drainage of Head & NeckRoopa PatilNo ratings yet
- Treating Breast Cancer: Local TreatmentsDocument99 pagesTreating Breast Cancer: Local Treatmentssuci triana putriNo ratings yet
- Anatomy and Physiology of Lymphatic SystemDocument24 pagesAnatomy and Physiology of Lymphatic Systemamber tariqNo ratings yet
- LYMPHOMADocument12 pagesLYMPHOMAश्रीकृष्ण हेङ्गजूNo ratings yet
- Lymphatic SystemDocument22 pagesLymphatic SystemdrynwhylNo ratings yet
- Examination of Lymph Nodes of Head and NeckDocument58 pagesExamination of Lymph Nodes of Head and NeckabdulrazaqNo ratings yet
- Micronatomy pt2Document35 pagesMicronatomy pt2poopziNo ratings yet
- PHC411 - Practical 3 - Oct 2021 - StudentsDocument14 pagesPHC411 - Practical 3 - Oct 2021 - StudentsALISYA SOPHIA MOHAMMAD ABU SHAHID CHRISNo ratings yet
- Advanced Piezoelectric Materials Science and Technology 2nd Edition Kenji Uchino Full ChapterDocument24 pagesAdvanced Piezoelectric Materials Science and Technology 2nd Edition Kenji Uchino Full Chapterwejzagough100% (3)
- 24 Lecture PresentationDocument85 pages24 Lecture PresentationWide PutinNo ratings yet
- Abreastcancerstory Breastcancertreatmentreview 100624205948 Phpapp01Document552 pagesAbreastcancerstory Breastcancertreatmentreview 100624205948 Phpapp01muhammad yamin100% (1)
- The SPLENOMEGALY Power Point Dr. CosmescuDocument29 pagesThe SPLENOMEGALY Power Point Dr. CosmescuNisha PillaiNo ratings yet
- Crystal Shipping, Inc. vs. Natividad, 473 SCRA 559 (2005Document15 pagesCrystal Shipping, Inc. vs. Natividad, 473 SCRA 559 (2005Mei SuNo ratings yet
- HistologyDocument18 pagesHistologyGlavinorNo ratings yet
- Immunology Tonsil 2Document31 pagesImmunology Tonsil 2ariienndrrahanniie100% (1)
- Leadership Initiative To Enrich The Hiv/aids Education For Senior High SchoolsDocument21 pagesLeadership Initiative To Enrich The Hiv/aids Education For Senior High SchoolsIshika JainNo ratings yet
- Ultrasound of Thyroid, Parathyroid Glands and Neck Lymph Nodes - SolbiatiDocument14 pagesUltrasound of Thyroid, Parathyroid Glands and Neck Lymph Nodes - Solbiatidrmoscalin8774No ratings yet
- Immune System Overview - Understanding The Immune SystemDocument60 pagesImmune System Overview - Understanding The Immune Systemlourdesbr100% (2)
- LimfadenitisDocument24 pagesLimfadenitisrahmah ningsihNo ratings yet
- Лекция 5Document27 pagesЛекция 5Oriyomi OdekunleNo ratings yet
- 4 2Document3 pages4 2Stefia AisyahNo ratings yet
- LyphomaDocument7 pagesLyphomaNambuye Midyero AhmedNo ratings yet
- 1 Крок 1 ЛС KROK 2007 2017 English BiochemistryDocument480 pages1 Крок 1 ЛС KROK 2007 2017 English BiochemistryAbhani MøhitNo ratings yet
- How I Evaluate The Neck Lymph NodesDocument29 pagesHow I Evaluate The Neck Lymph NodesMartín Avellaneda SantoméNo ratings yet
- 12th Zoology Practical Study Material English MediumDocument24 pages12th Zoology Practical Study Material English MediumKarthika KarthikaNo ratings yet
- Microscopic Descriptive TechniquesDocument13 pagesMicroscopic Descriptive TechniquesJill PNo ratings yet
- Clinical Feature of Lymphnode EnlargmentDocument47 pagesClinical Feature of Lymphnode Enlargmentjenny girlNo ratings yet
- Non HodgkinsDocument2 pagesNon HodgkinstapNo ratings yet
- Laboratory Methodsin ImmnunologyDocument58 pagesLaboratory Methodsin Immnunologyadi pNo ratings yet