Download as pdf or txt
You might also like
- Ant Architecture: The Wonder, Beauty, and Science of Underground NestsFrom EverandAnt Architecture: The Wonder, Beauty, and Science of Underground NestsNo ratings yet
- Chapter One 1.1 History and Meaning of SIWESDocument12 pagesChapter One 1.1 History and Meaning of SIWESDahiru Ibrahim100% (1)
- Secrets of The AnnunakiDocument17 pagesSecrets of The AnnunakiPietro Angelo Maria Massimi100% (1)
- Vol 34 No 1 PDFDocument56 pagesVol 34 No 1 PDFrlamtilaaNo ratings yet
- Encounters With Animal Minds: Journal of Consciousness StudiesDocument17 pagesEncounters With Animal Minds: Journal of Consciousness StudiesmanuelNo ratings yet
- Zebra SpiderDocument101 pagesZebra Spideralex100% (1)
- Mstudy Student's Guide For Topic TalkDocument30 pagesMstudy Student's Guide For Topic TalkArle CambalNo ratings yet
- Sample MonographDocument4 pagesSample MonographBucky_BallNo ratings yet
- Stem Level 1 Activity Form 1Document2 pagesStem Level 1 Activity Form 1api-668342729No ratings yet
- The Cultural Interface of Islander and Scientific KnowledgeDocument6 pagesThe Cultural Interface of Islander and Scientific KnowledgeFunny_PersonNo ratings yet
- Sample Monograph FinalDocument4 pagesSample Monograph FinalBucky_Ball0% (3)
- Exploration SpidersDocument4 pagesExploration SpidersEryNo ratings yet
- Meulenbelt, Hans 2016 Porophyllum Saxifrages Riddles The Saxifrage Magazine Vol 24 Pag 46-52Document7 pagesMeulenbelt, Hans 2016 Porophyllum Saxifrages Riddles The Saxifrage Magazine Vol 24 Pag 46-52Hans MeulenbeltNo ratings yet
- Amazing Amphibians: 30 Activities and Observations for Exploring Frogs, Toads, Salamanders, and MoreFrom EverandAmazing Amphibians: 30 Activities and Observations for Exploring Frogs, Toads, Salamanders, and MoreNo ratings yet
- Awesome Snake Science!: 40 Activities for Learning About SnakesFrom EverandAwesome Snake Science!: 40 Activities for Learning About SnakesRating: 5 out of 5 stars5/5 (1)
- NEL Portal Activity Idea Flora and Fauna All About Cassowaries PCF Braddell HeightsDocument8 pagesNEL Portal Activity Idea Flora and Fauna All About Cassowaries PCF Braddell HeightsLucas TobingNo ratings yet
- Stories in Tracks & Sign: Reading the Clues that Animals Leave BehindFrom EverandStories in Tracks & Sign: Reading the Clues that Animals Leave BehindRating: 3 out of 5 stars3/5 (3)
- Reading LP Catch Up FridayDocument4 pagesReading LP Catch Up Fridaymarivic.rosario005No ratings yet
- The Past Beneath Our Feet-Sj l3 May 2016Document5 pagesThe Past Beneath Our Feet-Sj l3 May 2016api-325718693No ratings yet
- CT Nature SpidersDocument1 pageCT Nature SpidersHartford CourantNo ratings yet
- September-October 2009 Coulee Birder Newsletter Coulee Region Audubon SocietyDocument4 pagesSeptember-October 2009 Coulee Birder Newsletter Coulee Region Audubon SocietyCoulee Region Audubon SocietyNo ratings yet
- France Plants As Inventors 1923 PDFDocument72 pagesFrance Plants As Inventors 1923 PDFpatriciaNo ratings yet
- Mocomi TimePass The Magazine - Issue 42Document15 pagesMocomi TimePass The Magazine - Issue 42Mocomi KidsNo ratings yet
- Convergent Evolution:: Cassin's Finches, An Irruptive Species?Document8 pagesConvergent Evolution:: Cassin's Finches, An Irruptive Species?PrescottNo ratings yet
- In Search of The UnknownDocument245 pagesIn Search of The UnknownunknownNo ratings yet
- Hatchet Unit 3Document11 pagesHatchet Unit 3Jamal Al-deenNo ratings yet
- A Conversation With William Rathje-Anthropology TodayDocument7 pagesA Conversation With William Rathje-Anthropology TodayAlexandra BejaranoNo ratings yet
- Week 7 - 2nd QuarterDocument2 pagesWeek 7 - 2nd QuarterMhai AdonaNo ratings yet
- Mocomi TimePass The Magazine - Issue 32Document14 pagesMocomi TimePass The Magazine - Issue 32Mocomi KidsNo ratings yet
- Exhibit-A: May 2020Document9 pagesExhibit-A: May 2020AnveshaNo ratings yet
- Wingmakers in The MakingDocument5 pagesWingmakers in The MakingigorfauxhawkNo ratings yet
- One Beetle Too Many by Kathryn Lasky - Q&A With AuthorDocument2 pagesOne Beetle Too Many by Kathryn Lasky - Q&A With AuthorCandlewick Press0% (1)
- 11 QitymaqoDocument3 pages11 Qitymaqopourya mashmoulNo ratings yet
- Alcock Cap 1Document12 pagesAlcock Cap 1Lilian Cerri MazzaNo ratings yet
- Alcock Cap 1Document12 pagesAlcock Cap 1Lilian Cerri MazzaNo ratings yet
- HorizonsDocument4 pagesHorizonsjenniferNo ratings yet
- LPCatchUpFriday Jan12 2024xDocument3 pagesLPCatchUpFriday Jan12 2024xliezle marie almadenNo ratings yet
- Dinosaurs Learn-AnywhereDocument31 pagesDinosaurs Learn-Anywhereana caetanoNo ratings yet
- My Gentle Barn by Ellie Laks - ExcerptDocument21 pagesMy Gentle Barn by Ellie Laks - ExcerptCrown Publishing Group100% (2)
- Solution Manual For Campbell Biology in Focus AP Edition 2nd by UrryDocument34 pagesSolution Manual For Campbell Biology in Focus AP Edition 2nd by Urrymosesfortread7gks6k100% (49)
- TOS Crow VocalizationsDocument2 pagesTOS Crow VocalizationsSyifa NathaniaNo ratings yet
- DocumentoDocument43 pagesDocumentoSofy SinchigaloNo ratings yet
- Autoethnography and Research 1Document4 pagesAutoethnography and Research 1api-710933460No ratings yet
- A Naturalist at Large: The Best Essays of Bernd HeinrichFrom EverandA Naturalist at Large: The Best Essays of Bernd HeinrichRating: 4.5 out of 5 stars4.5/5 (8)
- Jameson y Ratcliffe_2002 (Introducción a Scarabaeoidea)Document6 pagesJameson y Ratcliffe_2002 (Introducción a Scarabaeoidea)Carlos A. JulioNo ratings yet
- Scarabs 78Document22 pagesScarabs 78Carlos A. JulioNo ratings yet
- Wilson, E.O. 1987. The Little Things That Run The WorldDocument4 pagesWilson, E.O. 1987. The Little Things That Run The WorldCarlos A. JulioNo ratings yet
- Scarabs: Scarab Beetles at The Montréal InsectariumDocument22 pagesScarabs: Scarab Beetles at The Montréal InsectariumCarlos A. JulioNo ratings yet
- Scarabs: Regional Variation of Dorcus Titanus Cas-Tera: Lucanidae) in Northern and Southern South KoreaDocument18 pagesScarabs: Regional Variation of Dorcus Titanus Cas-Tera: Lucanidae) in Northern and Southern South KoreaCarlos A. JulioNo ratings yet
- Scarabs: Print ISSN 1937-8343 Online ISSN 1937-8351 Occasional Issue Number 77 May, 2015 LdmorokiicuggwvkqakatifwDocument26 pagesScarabs: Print ISSN 1937-8343 Online ISSN 1937-8351 Occasional Issue Number 77 May, 2015 LdmorokiicuggwvkqakatifwCarlos A. JulioNo ratings yet
- Arrow, 1944. Systematic Notes On Melolonthine Beetles Belonging To Holotrichia - Melolonthinae PDFDocument20 pagesArrow, 1944. Systematic Notes On Melolonthine Beetles Belonging To Holotrichia - Melolonthinae PDFCarlos A. JulioNo ratings yet
- Akakibara, A. M., & Evangelista, O. 2010. Membracis Foliata (Homoptera, Membracidae) and Allied SpeciesDocument20 pagesAkakibara, A. M., & Evangelista, O. 2010. Membracis Foliata (Homoptera, Membracidae) and Allied SpeciesCarlos A. JulioNo ratings yet
- Arrow G.J. 1946. A New Giant Species of Ruteline Coleoptera. (Rutelinae)Document2 pagesArrow G.J. 1946. A New Giant Species of Ruteline Coleoptera. (Rutelinae)Carlos A. JulioNo ratings yet
- AnAM L., 1994 - A Check-List of The Hungarian Scarabaeoidea With The Description of Ten New PDFDocument14 pagesAnAM L., 1994 - A Check-List of The Hungarian Scarabaeoidea With The Description of Ten New PDFCarlos A. JulioNo ratings yet
- 2006 ACDL-64 Melolonthidae Guerrero PDFDocument9 pages2006 ACDL-64 Melolonthidae Guerrero PDFCarlos A. JulioNo ratings yet
- Yan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaDocument12 pagesYan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaCarlos A. JulioNo ratings yet
- 2006 ACDL-65 Onthophagus Yucatanus PDFDocument4 pages2006 ACDL-65 Onthophagus Yucatanus PDFCarlos A. JulioNo ratings yet
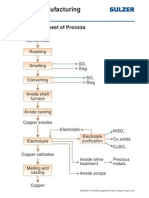
- Copper Manufacturing Process: General Flowsheet of ProcessDocument28 pagesCopper Manufacturing Process: General Flowsheet of ProcessflaviosazevedoNo ratings yet
- Geothermal DrillingDocument14 pagesGeothermal DrillingDaniel TobingNo ratings yet
- Newly Hired Teachers Lived Experiences in Classroom Management During The Full Face-to-Face Classes in The New Normal: A PhenomenologyDocument9 pagesNewly Hired Teachers Lived Experiences in Classroom Management During The Full Face-to-Face Classes in The New Normal: A PhenomenologyPsychology and Education: A Multidisciplinary JournalNo ratings yet
- Druid Spell Sheet - by John Bard (Printable)Document1 pageDruid Spell Sheet - by John Bard (Printable)Gabor PartizerNo ratings yet
- Tesol End Term AssignmentDocument7 pagesTesol End Term Assignmentakshita ramdasNo ratings yet
- Chatrapati Shivaji TurminusDocument6 pagesChatrapati Shivaji TurminusshrutiNo ratings yet
- PowerPact B-Frame Molded Case Circuit Breakers - BGA36020Document3 pagesPowerPact B-Frame Molded Case Circuit Breakers - BGA36020wajdyzNo ratings yet
- The Lovely BonesDocument2 pagesThe Lovely BonesJohn ConstantineNo ratings yet
- 1403 - Decoding The Education of Gifted Individual in The Ottoman Empire - Mimar Sinan's Education Case - İbrahim GÜLERDocument10 pages1403 - Decoding The Education of Gifted Individual in The Ottoman Empire - Mimar Sinan's Education Case - İbrahim GÜLERAHZeidanNo ratings yet
- Difference Between ASTM A435 and ASTM A578Document1 pageDifference Between ASTM A435 and ASTM A578Vijay BedakihaleNo ratings yet
- (379315304) 151221525-Deber-2Document19 pages(379315304) 151221525-Deber-2Daniel Armas100% (3)
- ADVANCE COPY 20230218 DILG ADVISORY No 02 On The LPADocument2 pagesADVANCE COPY 20230218 DILG ADVISORY No 02 On The LPABarangay MabuloNo ratings yet
- Holocaust Essay Thesis StatementDocument6 pagesHolocaust Essay Thesis Statementmmwsmltgg100% (2)
- Lesson Plan For DramaDocument6 pagesLesson Plan For DramaMerve MutluNo ratings yet
- Ghost Usb Honeypot MasterDocument15 pagesGhost Usb Honeypot MasterVinamra MittalNo ratings yet
- Violet Cough SyrupDocument16 pagesViolet Cough SyrupMrBond666No ratings yet
- Panasonic Air Condicioner CU-2E15LBE Service ManualDocument103 pagesPanasonic Air Condicioner CU-2E15LBE Service Manualjoxebike100% (1)
- Drum Dryers: Wan Ramli Wan DaudDocument15 pagesDrum Dryers: Wan Ramli Wan DaudYanuarRamadhanNo ratings yet
- ComercialInvoice PCW154986096Document1 pageComercialInvoice PCW154986096Quan VũNo ratings yet
- Job Circular For The Post of Manager, AE and SAEDocument2 pagesJob Circular For The Post of Manager, AE and SAEXYZNo ratings yet
- Belgium TemperingDocument14 pagesBelgium TemperingInes Kurfirst-VukušićNo ratings yet
- Proposed Arch Bridge On Lower Lake of Bhopal: A Case StudyDocument3 pagesProposed Arch Bridge On Lower Lake of Bhopal: A Case StudyInnovative Research PublicationsNo ratings yet
- Tutorial 1Document3 pagesTutorial 1Chong Cherng WongNo ratings yet
- Demosthenes, Speeches 1-17Document364 pagesDemosthenes, Speeches 1-17silviudx100% (1)
- Vega 450 Installation ManualDocument35 pagesVega 450 Installation ManualZeljko TomicNo ratings yet
- SAIL Challenges of Single Cell and Spatial ExperimentsDocument54 pagesSAIL Challenges of Single Cell and Spatial ExperimentsespartawithespartilhoNo ratings yet
- Quantity Estimate Store Room FinalDocument15 pagesQuantity Estimate Store Room FinalUjjwal ShresthaNo ratings yet
- 2020-COR-TXT Planetary Emergency 2-0-Aug2020Document9 pages2020-COR-TXT Planetary Emergency 2-0-Aug2020Mika DeverinNo ratings yet
- Badugu VaraKumar PLSQLDocument5 pagesBadugu VaraKumar PLSQLNavya DasariNo ratings yet