
Kap2 Enzimat
Kap2 Enzimat
You might also like
- Teza 1Document6 pagesTeza 1Swat AlbaniaNo ratings yet
- Neuro AnatomiaDocument80 pagesNeuro Anatomianina ninaNo ratings yet
- Anatomia Dhe Fiziologjia e NjeriutDocument12 pagesAnatomia Dhe Fiziologjia e Njeriutemi86de100% (4)
- GjjkhukjDocument17 pagesGjjkhukjAvni MehilliNo ratings yet
- Bazat e CitologjiseDocument14 pagesBazat e CitologjisePrince YTBNo ratings yet
- Proteinat - Strukturadhei FunkcioniDocument85 pagesProteinat - Strukturadhei FunkcioniFatlum Rushiti50% (2)
- Kapitulli 16Document7 pagesKapitulli 16Marina VrapiNo ratings yet
- 9 Proteinat Strukturadhei FunkcioniDocument85 pages9 Proteinat Strukturadhei FunkcioniUrim NdouNo ratings yet
- Funksionet e ADNDocument2 pagesFunksionet e ADNLorenc SakoNo ratings yet
- Pyetje Biologji-Matura Sh.Document24 pagesPyetje Biologji-Matura Sh.jasinNo ratings yet
- Biokimi PyetjeDocument10 pagesBiokimi PyetjeAsi SalajNo ratings yet
- 15 - GlikolizaDocument43 pages15 - GlikolizaFatlum RushitiNo ratings yet
- Biologji Variantib PDFDocument8 pagesBiologji Variantib PDFJoanna PoshnjaNo ratings yet
- Fiziologjia e Membranës Bërthamore.Document19 pagesFiziologjia e Membranës Bërthamore.Arbulena Morina100% (1)
- CitoplazmaDocument88 pagesCitoplazmaCoCo YT ffNo ratings yet
- Faktoret Etiologjik Lokal Dhe Te Pergjithshem Qe ShkaktojneDocument22 pagesFaktoret Etiologjik Lokal Dhe Te Pergjithshem Qe ShkaktojneArt AvdyliNo ratings yet
- EnzimatDocument9 pagesEnzimatRijadNo ratings yet
- Biologji e Thelluar ZgjidhjeDocument8 pagesBiologji e Thelluar ZgjidhjeOrjada100% (4)
- LatininishtDocument7 pagesLatininishtreni tofdazNo ratings yet
- Koncepte Bazë Rreth Qelizës (Rikujtesë!)Document581 pagesKoncepte Bazë Rreth Qelizës (Rikujtesë!)Meli HasaNo ratings yet
- Leksioni 12 - Aparati TretesDocument18 pagesLeksioni 12 - Aparati TretesDanjel Kola100% (1)
- StereokimiaDocument123 pagesStereokimiaartanNo ratings yet
- Zberthimi I Acideve YnderoreDocument2 pagesZberthimi I Acideve Ynderoreisuf fetaiNo ratings yet
- TretësiratDocument5 pagesTretësiratElda PernaskaNo ratings yet
- Teza Semundje Te Brendshme 2Document2 pagesTeza Semundje Te Brendshme 2klerida shehuNo ratings yet
- Tezat e BiologjiseDocument42 pagesTezat e BiologjiseAnisaNo ratings yet
- Projekt: VirusetDocument21 pagesProjekt: VirusetFero RigelaNo ratings yet
- Fiziologjia e SinapsitDocument6 pagesFiziologjia e SinapsitLulzim Bruçaj40% (5)
- Transporti I Materieve Nëpër Membranën QelizoreDocument3 pagesTransporti I Materieve Nëpër Membranën QelizorePhessyyNo ratings yet
- Gjenetik PyetjeDocument23 pagesGjenetik Pyetjealbin adiliNo ratings yet
- Punim Diplome F.RDocument38 pagesPunim Diplome F.ROLTA KASTRATINo ratings yet
- Cikli I KrebsitDocument14 pagesCikli I KrebsitBlodin Zylfiu100% (1)
- HISTOLOGJIDocument6 pagesHISTOLOGJIGent'No ratings yet
- EpigjenetikaDocument6 pagesEpigjenetikaArtanBajgoraNo ratings yet
- Test Pyetje Nga Biologjia 9.. Per Testin KombetarDocument20 pagesTest Pyetje Nga Biologjia 9.. Per Testin KombetarKosova ImeNo ratings yet
- Enzimat 3Document35 pagesEnzimat 3Arjan FejzaNo ratings yet
- Leksioni 3 Sistemet Koloidale, Difuzioni Dhe Presioni OsmotikDocument28 pagesLeksioni 3 Sistemet Koloidale, Difuzioni Dhe Presioni OsmotikFlorian PjetriNo ratings yet
- Hepatiti A B C D E GDocument75 pagesHepatiti A B C D E GlamzeqiriNo ratings yet
- Gjenetika MikrobikeDocument46 pagesGjenetika MikrobikeFero RigelaNo ratings yet
- BiokimiDocument6 pagesBiokimiBetta BeTtaNo ratings yet
- Leksioni 1, Zhvillimi Historik I Farmacise, StudentetDocument9 pagesLeksioni 1, Zhvillimi Historik I Farmacise, StudentetArber ZeqajNo ratings yet
- CikletDocument29 pagesCikletSpartak Beqiraj100% (1)
- Projekt BiologjiDocument27 pagesProjekt BiologjiRejdi Kacabumi100% (1)
- Biologji ThelluarDocument6 pagesBiologji ThelluarmigenaNo ratings yet
- Bazat e Infermieristikes - LegjislacioniDocument26 pagesBazat e Infermieristikes - LegjislacioniXhülîåVeselXhèñìNo ratings yet
- Fizike PDFDocument2 pagesFizike PDFxhulja50% (2)
- Semundjet Autosomale DominanteDocument18 pagesSemundjet Autosomale DominanteAjet AsaniNo ratings yet
- Tretësirat e Elektrolitëve - Hasan KaragjoziDocument25 pagesTretësirat e Elektrolitëve - Hasan KaragjoziLiridon SulejmaniNo ratings yet
- Biofizike Java e 1Document41 pagesBiofizike Java e 1Arif Margataj100% (1)
- Fizika XII - Grimcat Dhe Valet LUMTURI PEÇIDocument11 pagesFizika XII - Grimcat Dhe Valet LUMTURI PEÇIEnida MataNo ratings yet
- KarbohidratetDocument22 pagesKarbohidratetEdonisa100% (1)
- Biologji Sesioni 1Document6 pagesBiologji Sesioni 1Dorisa BahjaNo ratings yet
- Olimpiada Biologji ZgjidhjeDocument4 pagesOlimpiada Biologji ZgjidhjeXhesika PuskajNo ratings yet
- Grupet e GjakutDocument2 pagesGrupet e GjakutTafiiNo ratings yet
- HOMEOSTAZADocument17 pagesHOMEOSTAZANaza HondoziNo ratings yet
- TEZA Bio e ZgjidhurDocument59 pagesTEZA Bio e ZgjidhurKlajdi K LilaNo ratings yet
- Leksione 1Document121 pagesLeksione 1Kejsi MuhajNo ratings yet
- surenaleDocument12 pagessurenalenina ninaNo ratings yet
- tiroidjaDocument9 pagestiroidjanina ninaNo ratings yet
- Brecht PoeziDocument24 pagesBrecht Poezinina ninaNo ratings yet
- Hormonet - 1 - 5888Document47 pagesHormonet - 1 - 5888nina ninaNo ratings yet
- 9.3 Lipide - Lipoproteinat - 5014Document25 pages9.3 Lipide - Lipoproteinat - 5014nina ninaNo ratings yet
- Material Plotesues GlucidetdocDocument3 pagesMaterial Plotesues Glucidetdocnina ninaNo ratings yet
- 5.2 Cikli - I - Urese - 5488Document23 pages5.2 Cikli - I - Urese - 5488nina ninaNo ratings yet
- Perkusioni, PalpacioniDocument37 pagesPerkusioni, Palpacioninina ninaNo ratings yet
- 1.leksioni 1Document11 pages1.leksioni 1nina ninaNo ratings yet
- NukleotidetDocument2 pagesNukleotidetnina ninaNo ratings yet




















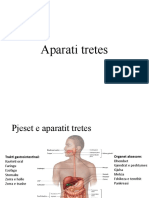
















































Download as pdf or txt
You might also like
- Teza 1Document6 pagesTeza 1Swat AlbaniaNo ratings yet
- Neuro AnatomiaDocument80 pagesNeuro Anatomianina ninaNo ratings yet
- Anatomia Dhe Fiziologjia e NjeriutDocument12 pagesAnatomia Dhe Fiziologjia e Njeriutemi86de100% (4)
- GjjkhukjDocument17 pagesGjjkhukjAvni MehilliNo ratings yet
- Bazat e CitologjiseDocument14 pagesBazat e CitologjisePrince YTBNo ratings yet
- Proteinat - Strukturadhei FunkcioniDocument85 pagesProteinat - Strukturadhei FunkcioniFatlum Rushiti50% (2)
- Kapitulli 16Document7 pagesKapitulli 16Marina VrapiNo ratings yet
- 9 Proteinat Strukturadhei FunkcioniDocument85 pages9 Proteinat Strukturadhei FunkcioniUrim NdouNo ratings yet
- Funksionet e ADNDocument2 pagesFunksionet e ADNLorenc SakoNo ratings yet
- Pyetje Biologji-Matura Sh.Document24 pagesPyetje Biologji-Matura Sh.jasinNo ratings yet
- Biokimi PyetjeDocument10 pagesBiokimi PyetjeAsi SalajNo ratings yet
- 15 - GlikolizaDocument43 pages15 - GlikolizaFatlum RushitiNo ratings yet
- Biologji Variantib PDFDocument8 pagesBiologji Variantib PDFJoanna PoshnjaNo ratings yet
- Fiziologjia e Membranës Bërthamore.Document19 pagesFiziologjia e Membranës Bërthamore.Arbulena Morina100% (1)
- CitoplazmaDocument88 pagesCitoplazmaCoCo YT ffNo ratings yet
- Faktoret Etiologjik Lokal Dhe Te Pergjithshem Qe ShkaktojneDocument22 pagesFaktoret Etiologjik Lokal Dhe Te Pergjithshem Qe ShkaktojneArt AvdyliNo ratings yet
- EnzimatDocument9 pagesEnzimatRijadNo ratings yet
- Biologji e Thelluar ZgjidhjeDocument8 pagesBiologji e Thelluar ZgjidhjeOrjada100% (4)
- LatininishtDocument7 pagesLatininishtreni tofdazNo ratings yet
- Koncepte Bazë Rreth Qelizës (Rikujtesë!)Document581 pagesKoncepte Bazë Rreth Qelizës (Rikujtesë!)Meli HasaNo ratings yet
- Leksioni 12 - Aparati TretesDocument18 pagesLeksioni 12 - Aparati TretesDanjel Kola100% (1)
- StereokimiaDocument123 pagesStereokimiaartanNo ratings yet
- Zberthimi I Acideve YnderoreDocument2 pagesZberthimi I Acideve Ynderoreisuf fetaiNo ratings yet
- TretësiratDocument5 pagesTretësiratElda PernaskaNo ratings yet
- Teza Semundje Te Brendshme 2Document2 pagesTeza Semundje Te Brendshme 2klerida shehuNo ratings yet
- Tezat e BiologjiseDocument42 pagesTezat e BiologjiseAnisaNo ratings yet
- Projekt: VirusetDocument21 pagesProjekt: VirusetFero RigelaNo ratings yet
- Fiziologjia e SinapsitDocument6 pagesFiziologjia e SinapsitLulzim Bruçaj40% (5)
- Transporti I Materieve Nëpër Membranën QelizoreDocument3 pagesTransporti I Materieve Nëpër Membranën QelizorePhessyyNo ratings yet
- Gjenetik PyetjeDocument23 pagesGjenetik Pyetjealbin adiliNo ratings yet
- Punim Diplome F.RDocument38 pagesPunim Diplome F.ROLTA KASTRATINo ratings yet
- Cikli I KrebsitDocument14 pagesCikli I KrebsitBlodin Zylfiu100% (1)
- HISTOLOGJIDocument6 pagesHISTOLOGJIGent'No ratings yet
- EpigjenetikaDocument6 pagesEpigjenetikaArtanBajgoraNo ratings yet
- Test Pyetje Nga Biologjia 9.. Per Testin KombetarDocument20 pagesTest Pyetje Nga Biologjia 9.. Per Testin KombetarKosova ImeNo ratings yet
- Enzimat 3Document35 pagesEnzimat 3Arjan FejzaNo ratings yet
- Leksioni 3 Sistemet Koloidale, Difuzioni Dhe Presioni OsmotikDocument28 pagesLeksioni 3 Sistemet Koloidale, Difuzioni Dhe Presioni OsmotikFlorian PjetriNo ratings yet
- Hepatiti A B C D E GDocument75 pagesHepatiti A B C D E GlamzeqiriNo ratings yet
- Gjenetika MikrobikeDocument46 pagesGjenetika MikrobikeFero RigelaNo ratings yet
- BiokimiDocument6 pagesBiokimiBetta BeTtaNo ratings yet
- Leksioni 1, Zhvillimi Historik I Farmacise, StudentetDocument9 pagesLeksioni 1, Zhvillimi Historik I Farmacise, StudentetArber ZeqajNo ratings yet
- CikletDocument29 pagesCikletSpartak Beqiraj100% (1)
- Projekt BiologjiDocument27 pagesProjekt BiologjiRejdi Kacabumi100% (1)
- Biologji ThelluarDocument6 pagesBiologji ThelluarmigenaNo ratings yet
- Bazat e Infermieristikes - LegjislacioniDocument26 pagesBazat e Infermieristikes - LegjislacioniXhülîåVeselXhèñìNo ratings yet
- Fizike PDFDocument2 pagesFizike PDFxhulja50% (2)
- Semundjet Autosomale DominanteDocument18 pagesSemundjet Autosomale DominanteAjet AsaniNo ratings yet
- Tretësirat e Elektrolitëve - Hasan KaragjoziDocument25 pagesTretësirat e Elektrolitëve - Hasan KaragjoziLiridon SulejmaniNo ratings yet
- Biofizike Java e 1Document41 pagesBiofizike Java e 1Arif Margataj100% (1)
- Fizika XII - Grimcat Dhe Valet LUMTURI PEÇIDocument11 pagesFizika XII - Grimcat Dhe Valet LUMTURI PEÇIEnida MataNo ratings yet
- KarbohidratetDocument22 pagesKarbohidratetEdonisa100% (1)
- Biologji Sesioni 1Document6 pagesBiologji Sesioni 1Dorisa BahjaNo ratings yet
- Olimpiada Biologji ZgjidhjeDocument4 pagesOlimpiada Biologji ZgjidhjeXhesika PuskajNo ratings yet
- Grupet e GjakutDocument2 pagesGrupet e GjakutTafiiNo ratings yet
- HOMEOSTAZADocument17 pagesHOMEOSTAZANaza HondoziNo ratings yet
- TEZA Bio e ZgjidhurDocument59 pagesTEZA Bio e ZgjidhurKlajdi K LilaNo ratings yet
- Leksione 1Document121 pagesLeksione 1Kejsi MuhajNo ratings yet
- surenaleDocument12 pagessurenalenina ninaNo ratings yet
- tiroidjaDocument9 pagestiroidjanina ninaNo ratings yet
- Brecht PoeziDocument24 pagesBrecht Poezinina ninaNo ratings yet
- Hormonet - 1 - 5888Document47 pagesHormonet - 1 - 5888nina ninaNo ratings yet
- 9.3 Lipide - Lipoproteinat - 5014Document25 pages9.3 Lipide - Lipoproteinat - 5014nina ninaNo ratings yet
- Material Plotesues GlucidetdocDocument3 pagesMaterial Plotesues Glucidetdocnina ninaNo ratings yet
- 5.2 Cikli - I - Urese - 5488Document23 pages5.2 Cikli - I - Urese - 5488nina ninaNo ratings yet
- Perkusioni, PalpacioniDocument37 pagesPerkusioni, Palpacioninina ninaNo ratings yet
- 1.leksioni 1Document11 pages1.leksioni 1nina ninaNo ratings yet
- NukleotidetDocument2 pagesNukleotidetnina ninaNo ratings yet