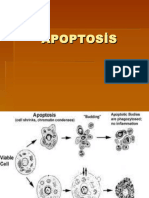
Download as pdf or txt
You might also like
- Ganong TürkçeDocument732 pagesGanong TürkçeCan Aras67% (9)
- ReprogramingDocument9 pagesReprogramingDr.Serkan SenerNo ratings yet
- Alzheimer Hastalığının Fizyopatolojisi WWW Stetuskop - Com 7Document10 pagesAlzheimer Hastalığının Fizyopatolojisi WWW Stetuskop - Com 7STETUSKOP.COM-TIP VE TUS'UN MERKEZİNo ratings yet
- 1 Araştirma Suat ÇakinaDocument5 pages1 Araştirma Suat ÇakinaJantikaNo ratings yet
- Ders 6 Nörodejeneratif HastalıklarDocument56 pagesDers 6 Nörodejeneratif HastalıklarNeslihan BOZKURTNo ratings yet
- Epilepsi GentkDocument9 pagesEpilepsi GentkyavuzNo ratings yet
- İmmun Kökenli Nöropatiler WWW Stetuskop - Com 6Document5 pagesİmmun Kökenli Nöropatiler WWW Stetuskop - Com 6STETUSKOP.COM-TIP VE TUS'UN MERKEZİNo ratings yet
- ALS (Amyotrofik Lateral Skleroz)Document26 pagesALS (Amyotrofik Lateral Skleroz)MedineNo ratings yet
- T.C. Sağlik Bilimleri Üniversitesi, Kayseri Şehir Eğitim Ve Araştirma Hastanesi Çocuk Sağliği Ve Hastaliklari KliniğiDocument57 pagesT.C. Sağlik Bilimleri Üniversitesi, Kayseri Şehir Eğitim Ve Araştirma Hastanesi Çocuk Sağliği Ve Hastaliklari KliniğipuhumightNo ratings yet
- EpigenetikDocument117 pagesEpigenetikEylül Nur ÇakırNo ratings yet
- Gelişim Genetiği 30.05.2011 (Epigenetik)Document116 pagesGelişim Genetiği 30.05.2011 (Epigenetik)zuhal100% (1)
- Psikiyatrik Elektrofizyoloji DemansDocument32 pagesPsikiyatrik Elektrofizyoloji DemansmkarikanNo ratings yet
- Apoptoz Ve Nekrozun Molekler Mekanizmas - (#29409) - 25371Document14 pagesApoptoz Ve Nekrozun Molekler Mekanizmas - (#29409) - 25371Hürkan AykaşNo ratings yet
- IM281818.. 010101 Ababsdb ,,,,, Ekwkwwj18 Ananqnwn ...... J-21-297-EnDocument4 pagesIM281818.. 010101 Ababsdb ,,,,, Ekwkwwj18 Ananqnwn ...... J-21-297-EnRed DiggerNo ratings yet
- Mikoloji 5Document31 pagesMikoloji 526ugurNo ratings yet
- Expression of Ki-67, Bcl-2 and Bax in The First Trimester Abortion MaterialsDocument7 pagesExpression of Ki-67, Bcl-2 and Bax in The First Trimester Abortion MaterialsSon GohanNo ratings yet
- Epi̇geneti̇k Ve Epi̇geneti̇k Mekani̇zmalarDocument70 pagesEpi̇geneti̇k Ve Epi̇geneti̇k Mekani̇zmalarCevikAlp75% (12)
- Hücre Zedelenmesi̇Document15 pagesHücre Zedelenmesi̇Mesut AydinNo ratings yet
- Eozifilik ÖzofajitDocument3 pagesEozifilik Özofajitabdullatif şirinNo ratings yet
- Models of Experimental Epilepsy 3193Document6 pagesModels of Experimental Epilepsy 3193elmin_pctNo ratings yet
- NutrigeneticDocument46 pagesNutrigeneticwhiteyasminNo ratings yet
- WNT-Beta-Katenin Sinyal Yolunun Sitoplazmik Biyomolek - Lleri (#63513) - 53537Document11 pagesWNT-Beta-Katenin Sinyal Yolunun Sitoplazmik Biyomolek - Lleri (#63513) - 53537Zeynep YazıcıoğluNo ratings yet
- Epigenetik MekanizmalarDocument34 pagesEpigenetik Mekanizmalarenes özNo ratings yet
- Üreme Yaşam Öyküsüne Odaklanan Evrimsel Uyumsuzluk Hastalığı Olarak Alzheimer HastalığıDocument7 pagesÜreme Yaşam Öyküsüne Odaklanan Evrimsel Uyumsuzluk Hastalığı Olarak Alzheimer HastalığıYusuf DumanNo ratings yet
- X Gamma GlobulinemiDocument12 pagesX Gamma GlobulinemiulsynnNo ratings yet
- File 1770 PDFDocument301 pagesFile 1770 PDFasdssaNo ratings yet
- Tipbil27 4 13Document7 pagesTipbil27 4 13Mustafa CingözNo ratings yet
- P53 Ve ApoptozDocument19 pagesP53 Ve ApoptozzuhalNo ratings yet
- Buzağılarda İmmunomodülatör Zylexis'in Kan Glutatyon, Malondialdehit, Nitrik Oksit, Toplam Ve Lipit Bağlı Sialik Asit Seviyelerine EtkileriDocument6 pagesBuzağılarda İmmunomodülatör Zylexis'in Kan Glutatyon, Malondialdehit, Nitrik Oksit, Toplam Ve Lipit Bağlı Sialik Asit Seviyelerine EtkileriCeyda AydınNo ratings yet
- Eritrosit SunusuDocument176 pagesEritrosit SunusuCan ArasNo ratings yet
- Otoimmun Kolanjit Gokhan TumgorDocument39 pagesOtoimmun Kolanjit Gokhan Tumgorfurkanfurkan105105No ratings yet
- EPİGENETİKDocument25 pagesEPİGENETİKedaguc85No ratings yet
- GBS Tedavi RehberiDocument15 pagesGBS Tedavi RehberiSüleyman TurabiNo ratings yet
- Bolum3Document31 pagesBolum3Zae M HakimNo ratings yet
- Histoloji Ve Embiryoloji SunumDocument31 pagesHistoloji Ve Embiryoloji Sunumpolatseda159No ratings yet
- Akut Lenfoid Lösemi (ALL) - DR - Adem - KutanisDocument70 pagesAkut Lenfoid Lösemi (ALL) - DR - Adem - KutanisGercekhNo ratings yet
- 693242Document73 pages693242Emir ŞenNo ratings yet
- VarikoselDocument9 pagesVarikoselsifirzeroNo ratings yet
- Motor Nöron HastaliğiDocument6 pagesMotor Nöron HastaliğiŞükrü BoybayNo ratings yet
- 10 30569 Adiyamansaglik 396211 446842Document19 pages10 30569 Adiyamansaglik 396211 446842Ujjawal KumarNo ratings yet
- Hepatik Ensefalopati: Mehmet Fatih ErbayDocument6 pagesHepatik Ensefalopati: Mehmet Fatih ErbayCici PatresiaNo ratings yet
- TB HormonlarDocument41 pagesTB HormonlarrubNo ratings yet
- Eşi̇ Sma Taşiyicisi Olan Ki̇şi̇ İçi̇n Bi̇lgi̇ BeyaniDocument2 pagesEşi̇ Sma Taşiyicisi Olan Ki̇şi̇ İçi̇n Bi̇lgi̇ BeyaniLazgin DemirNo ratings yet
- 10 31832-smj 1066524-2226223Document14 pages10 31832-smj 1066524-2226223SAMET DÖNMEZNo ratings yet
- Genetiğe Giriş1-Öğrenci - 1 Ve 2 KonuDocument31 pagesGenetiğe Giriş1-Öğrenci - 1 Ve 2 KonuPınar ÖzdemirNo ratings yet
- Hücre Yaşlanması - SonDocument37 pagesHücre Yaşlanması - SonBerkay DncNo ratings yet
- 274563Document64 pages274563Göksu BakiNo ratings yet
- Alzheimer Hastal - Ve Koruyucu Besin - Eleri (#258507) - 224974 PDFDocument7 pagesAlzheimer Hastal - Ve Koruyucu Besin - Eleri (#258507) - 224974 PDFAnonymous IGYH26sky3No ratings yet
- Fizyopatoloji GirişDocument36 pagesFizyopatoloji GirişsuedaizemNo ratings yet
- PolimorfizmDocument31 pagesPolimorfizmenes özNo ratings yet
- Nonfonksi̇yonel Adrenal İnsi̇dentalomali Hastalarda Serum Vi̇sfati̇n Düzeyi̇ İle Kardi̇yometaboli̇k Ri̇sk Faktörleri̇Document51 pagesNonfonksi̇yonel Adrenal İnsi̇dentalomali Hastalarda Serum Vi̇sfati̇n Düzeyi̇ İle Kardi̇yometaboli̇k Ri̇sk Faktörleri̇kimmyNo ratings yet
- Kli̇ni̇k Enzi̇moloji̇Document4 pagesKli̇ni̇k Enzi̇moloji̇NAME SURNAMENo ratings yet
- Belge 1Document11 pagesBelge 1Deniz KiziltanNo ratings yet
- Programlanmýþ Hücre Ölümü: Apopitozis: Apopitosis: Programmed Cell DeathDocument5 pagesProgramlanmýþ Hücre Ölümü: Apopitozis: Apopitosis: Programmed Cell Deathcelal yamanNo ratings yet
- Kolorektal Karsi̇nomlarda Adezyon Molekülleri̇ Ve Anji̇ogenezi̇n Kli̇ni̇kopatoloji̇k Parametrelerle İli̇şki̇leri̇Document92 pagesKolorektal Karsi̇nomlarda Adezyon Molekülleri̇ Ve Anji̇ogenezi̇n Kli̇ni̇kopatoloji̇k Parametrelerle İli̇şki̇leri̇SelinNo ratings yet
- Şizofreni DönerkapıDocument87 pagesŞizofreni DönerkapıSedat KayaNo ratings yet
- Genel Patoloji - 01 02.hafta Dejenerasyon - NekrozDocument249 pagesGenel Patoloji - 01 02.hafta Dejenerasyon - NekrozCeren CelayirNo ratings yet
- K - Peklerde Pyometran - N Tan - S - Nda Laboratuar, Ultrasonografi Ve Vaginal Sitoloji... (#163752) - 144597Document8 pagesK - Peklerde Pyometran - N Tan - S - Nda Laboratuar, Ultrasonografi Ve Vaginal Sitoloji... (#163752) - 144597Şahin ŞahinNo ratings yet
- 10 Geneti̇k Çeşi̇tli̇li̇ği̇n (Geneti̇k Varyasyonlarin) Nedenleri̇Document1 page10 Geneti̇k Çeşi̇tli̇li̇ği̇n (Geneti̇k Varyasyonlarin) Nedenleri̇esmanurozer1142No ratings yet
- International Researches in Health and Natural SciencesDocument117 pagesInternational Researches in Health and Natural Sciencesşenol topuzNo ratings yet
- 1822 Yanlish - Yonde - Kuantum - Sichramalar Charles - M.Wynn Arthur - W.Wiggins Chev Ayqut - Kence 2005 229sDocument229 pages1822 Yanlish - Yonde - Kuantum - Sichramalar Charles - M.Wynn Arthur - W.Wiggins Chev Ayqut - Kence 2005 229sşenol topuzNo ratings yet
- Ünite 3 - Algı, Tutum, AtıfDocument85 pagesÜnite 3 - Algı, Tutum, Atıfşenol topuzNo ratings yet
- Ünite 1 - Davranış Bilimine GirişDocument22 pagesÜnite 1 - Davranış Bilimine Girişşenol topuzNo ratings yet
- Anti̇korlar Ve Kullanim AlanlariDocument76 pagesAnti̇korlar Ve Kullanim Alanlarişenol topuzNo ratings yet