Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Project Report For Tomato Processing PlantDocument2 pagesProject Report For Tomato Processing PlantRajib Deb100% (2)
- BKM Z60 80 B III Service Manual PDFDocument54 pagesBKM Z60 80 B III Service Manual PDFKelly VillamilNo ratings yet
- A Comparison of Immunohistochemical and Silver Staining Methods For The Detection of Diffuse Plaques ... (2006)Document13 pagesA Comparison of Immunohistochemical and Silver Staining Methods For The Detection of Diffuse Plaques ... (2006)Apróné Török IbolyaNo ratings yet
- Health Regulation of Dietary SupplementsDocument22 pagesHealth Regulation of Dietary SupplementsApróné Török IbolyaNo ratings yet
- Intrinsic and Extrinsic Wiring of CA3 - Indications For Connectional Heterogeneity (2007)Document10 pagesIntrinsic and Extrinsic Wiring of CA3 - Indications For Connectional Heterogeneity (2007)Apróné Török IbolyaNo ratings yet
- Inhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Document13 pagesInhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Apróné Török IbolyaNo ratings yet
- The Basal Forebrain Cholinergic Projection System in Mice - Chapter 28Document35 pagesThe Basal Forebrain Cholinergic Projection System in Mice - Chapter 28Apróné Török IbolyaNo ratings yet
- Neto1 and Neto2 - Auxiliary Subunits That Determine Key Properties of Native Kainate Receptors (2012)Document7 pagesNeto1 and Neto2 - Auxiliary Subunits That Determine Key Properties of Native Kainate Receptors (2012)Apróné Török IbolyaNo ratings yet
- Cell Signalling Biology - Ion Channels (2012)Document72 pagesCell Signalling Biology - Ion Channels (2012)Apróné Török IbolyaNo ratings yet
- Natural Products For Neurodegenerative Diseases (Neurosignals) (2005)Document84 pagesNatural Products For Neurodegenerative Diseases (Neurosignals) (2005)Apróné Török IbolyaNo ratings yet
- Biochemical and Biophysical Research Communications: Takafumi Ohtsuka, Masato Yano, Hideyuki OkanoDocument6 pagesBiochemical and Biophysical Research Communications: Takafumi Ohtsuka, Masato Yano, Hideyuki OkanoApróné Török IbolyaNo ratings yet
- Acute and Chronic Responses To The Convulsant Pilocarpine in DBA2J and AJ Mice (2007)Document11 pagesAcute and Chronic Responses To The Convulsant Pilocarpine in DBA2J and AJ Mice (2007)Apróné Török IbolyaNo ratings yet
- Vagus Nerve Stimulation and Magnet Use Optimizing Benefits (2009)Document4 pagesVagus Nerve Stimulation and Magnet Use Optimizing Benefits (2009)Apróné Török IbolyaNo ratings yet
- Vagus Nerve Stimulation and Magnet Use - Optimizing Benefits (2009)Document4 pagesVagus Nerve Stimulation and Magnet Use - Optimizing Benefits (2009)Apróné Török IbolyaNo ratings yet
- IOCLDocument94 pagesIOCLDebabrat SaikiaNo ratings yet
- Emotional Disturbance Annotated BibliographyDocument3 pagesEmotional Disturbance Annotated Bibliographyapi-335203912No ratings yet
- Y2 Series: Three Phase Cast Iron MotorDocument4 pagesY2 Series: Three Phase Cast Iron MotorMark Christian TorresNo ratings yet
- TR MassageTherapy NCIIDocument91 pagesTR MassageTherapy NCIIRebu Leno Rnlptlmtcssi100% (2)
- Nursing Care Plan For A Person With Croup Nursing DiagnosisDocument2 pagesNursing Care Plan For A Person With Croup Nursing DiagnosisMonica Rivera100% (1)
- Codein 15 MG Aptil - Mei 2023Document4 pagesCodein 15 MG Aptil - Mei 2023Nikolas EdwinNo ratings yet
- 273 Pocket Theatre Training ActivityDocument1 page273 Pocket Theatre Training Activityxxxxxx100% (1)
- Contraception Your Questions AnsweredDocument144 pagesContraception Your Questions AnsweredMohamed SaeedNo ratings yet
- Body ImageDocument1 pageBody Imageletitia2hintonNo ratings yet
- The Foundations of ChemistryDocument38 pagesThe Foundations of ChemistryJesusBlasVitangcolNo ratings yet
- Near-Infrared Spectroscopy Measurements of Cerebral Oxygenation in Newborns During Immediate Postnatal AdaptationDocument6 pagesNear-Infrared Spectroscopy Measurements of Cerebral Oxygenation in Newborns During Immediate Postnatal Adaptationcodruta_holtzNo ratings yet
- Roastilino Manual v2 1 21 6 13Document9 pagesRoastilino Manual v2 1 21 6 13Bigman74066No ratings yet
- Human Rights & Status of Woman in IslamDocument7 pagesHuman Rights & Status of Woman in Islamsweetmano100% (1)
- Acute Lymphoblastic Leukemia RadiopediaDocument2 pagesAcute Lymphoblastic Leukemia RadiopediaRani Henty NovitaNo ratings yet
- Polyhydraminos and OligohydraminosDocument11 pagesPolyhydraminos and OligohydraminosMelissa Catherine ChinNo ratings yet
- Nippon Wall Putty A-100: Water BasedDocument3 pagesNippon Wall Putty A-100: Water BasedNippon Paint PakistanNo ratings yet
- 2020-06-18 Calvert County TimesDocument32 pages2020-06-18 Calvert County TimesSouthern Maryland OnlineNo ratings yet
- Panel Board Diagram Fluorescent Light Fixture Mounting DetailDocument1 pagePanel Board Diagram Fluorescent Light Fixture Mounting DetailWilbert Reuyan100% (1)
- ASSESSOR ScriptDocument3 pagesASSESSOR ScriptJonalyn Lajera BasayaNo ratings yet
- Dheeraj Coca Cola MbaDocument84 pagesDheeraj Coca Cola Mbaranaindia2011100% (1)
- Shibu George Et Al. - 2023Document16 pagesShibu George Et Al. - 2023apatzNo ratings yet
- Lesson Plan in Top of Stove Cooking Equipment and Baking EquipmentDocument16 pagesLesson Plan in Top of Stove Cooking Equipment and Baking EquipmentNiña Mae SabilloNo ratings yet
- HomoeopathyDocument354 pagesHomoeopathyEla Drzewiecka100% (4)
- Btech Me 5 Sem Heat and Mass Transfer Eme504 2020Document2 pagesBtech Me 5 Sem Heat and Mass Transfer Eme504 2020SuryaNo ratings yet
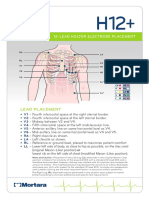
- ECG Holter H12+Document2 pagesECG Holter H12+Dhilah 9246No ratings yet
- Rili DLP MutationDocument16 pagesRili DLP MutationJohn Bernard RiliNo ratings yet
- Term Paper On Organic FarmingDocument8 pagesTerm Paper On Organic Farmingc5rm5y5p100% (1)
- Dance TherapyDocument1 pageDance TherapyRoxanne Dooma0% (1)