
Dandan Li, Liangkun Long Dan Shaojun Ding. 2020. Alkaline Organosolv Pretreatment of Different
Dandan Li, Liangkun Long Dan Shaojun Ding. 2020. Alkaline Organosolv Pretreatment of Different
You might also like
- Alkaline Organosolv Pretreatment of Different Sorghum Stem Parts For Enhancing The Total Reducing Sugar Yields and P-Coumaric Acid ReleaseDocument14 pagesAlkaline Organosolv Pretreatment of Different Sorghum Stem Parts For Enhancing The Total Reducing Sugar Yields and P-Coumaric Acid Releaseuliseshbq7No ratings yet
- Industrially Relevant Hydrolyzability and Fermentability of Sugarcane Bagasse Improved Effectively by Glycerol Organosolv PretreatmentDocument13 pagesIndustrially Relevant Hydrolyzability and Fermentability of Sugarcane Bagasse Improved Effectively by Glycerol Organosolv PretreatmentHarshad Vinay SavantNo ratings yet
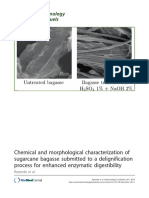
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- A Novel Recyclable Furoic Acid-Assisted Pretreatment For Sugarcane Bagasse Biorefinery in Co-Production of Xylooligosaccharides and GlucoseDocument8 pagesA Novel Recyclable Furoic Acid-Assisted Pretreatment For Sugarcane Bagasse Biorefinery in Co-Production of Xylooligosaccharides and GlucoseSthiden Sanabria ChaconNo ratings yet
- Feasibility Study of On-Site Solid-State Enzyme ProductionDocument15 pagesFeasibility Study of On-Site Solid-State Enzyme ProductionOuti ElinaNo ratings yet
- Influence of Sulfuric Acid Pretreatment and Inhibitor of Sugarcane Bagasse On The Production of Fermentable Sugar and EthanolDocument13 pagesInfluence of Sulfuric Acid Pretreatment and Inhibitor of Sugarcane Bagasse On The Production of Fermentable Sugar and EthanolJesusPJNo ratings yet
- Integrating Sugarcane Molasses Into Sequential Cellulosic Biofuel Production Based On SSF Process of High Solid LoadingDocument9 pagesIntegrating Sugarcane Molasses Into Sequential Cellulosic Biofuel Production Based On SSF Process of High Solid LoadingSamuel GamorNo ratings yet
- An Innovative Approach of Mixed Enzymatic Venture For 2G Ethanol Production From Lignocellulosic Feedstock PDFDocument10 pagesAn Innovative Approach of Mixed Enzymatic Venture For 2G Ethanol Production From Lignocellulosic Feedstock PDFRajiv Chandra RajakNo ratings yet
- International Journal of Biological Macromolecules Comparative Study 1-S2.0-S0141813019393699-MainDocument9 pagesInternational Journal of Biological Macromolecules Comparative Study 1-S2.0-S0141813019393699-MainOmar KHELILNo ratings yet
- Simultaneous Enzymatic Saccharification and Comminution For The Valorization of Lignocellulosic Biomass Toward Natural ProductsDocument11 pagesSimultaneous Enzymatic Saccharification and Comminution For The Valorization of Lignocellulosic Biomass Toward Natural ProductsSachin SharmaNo ratings yet
- Tongco2020 Article EnhancementOfHydrolysisAndBiogDocument9 pagesTongco2020 Article EnhancementOfHydrolysisAndBiogJim TsikasNo ratings yet
- Improvement of Enzymatic SaccharificationDocument11 pagesImprovement of Enzymatic SaccharificationpepandradeNo ratings yet
- International Journal of Biological MacromoleculesDocument7 pagesInternational Journal of Biological MacromoleculesEzanaNo ratings yet
- 14043-Article Text PDF-58576-2-10-20190902Document8 pages14043-Article Text PDF-58576-2-10-20190902R. VersteeghNo ratings yet
- 1 s2.0 S0360319922042458 MainDocument11 pages1 s2.0 S0360319922042458 Mainranjani093No ratings yet
- Isolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilDocument8 pagesIsolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilATSNo ratings yet
- Enzymatic Digestibility of Pretreated Dewaxed Bamboo Residues As Feedstock For Bioethanol ProductionDocument7 pagesEnzymatic Digestibility of Pretreated Dewaxed Bamboo Residues As Feedstock For Bioethanol ProductionKevin Hernandez Duque100% (1)
- Materials: Multi-Functional Laccase Immobilized Hydrogel Microparticles For Efficient Removal of Bisphenol ADocument14 pagesMaterials: Multi-Functional Laccase Immobilized Hydrogel Microparticles For Efficient Removal of Bisphenol AcarolasbdNo ratings yet
- Accepted Manuscript: Bioresource TechnologyDocument32 pagesAccepted Manuscript: Bioresource TechnologyJulio MoralesNo ratings yet
- Structured Polyvinyl AlcoholZeoliteCarbon CompositDocument11 pagesStructured Polyvinyl AlcoholZeoliteCarbon CompositFarah TalibNo ratings yet
- Biochemical Characterization and SynergismDocument12 pagesBiochemical Characterization and SynergismJayesh PatidarNo ratings yet
- Biomass and Bioenergy: Research PaperDocument10 pagesBiomass and Bioenergy: Research PaperPaulina Lpz JrzNo ratings yet
- Sanjeetha Assignment 3 Proposal ReportDocument20 pagesSanjeetha Assignment 3 Proposal ReportSanjith KaurNo ratings yet
- 11403-Article Text PDF-34245-3-10-20180516Document12 pages11403-Article Text PDF-34245-3-10-20180516Mariana CampoNo ratings yet
- Dye RemovalDocument12 pagesDye RemovalaisyahNo ratings yet
- Research Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionDocument10 pagesResearch Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionShankar AcharNo ratings yet
- Characterization of Cell Wall Components of Wheat Bran Following Hydrothermal Pretreatment and FractionationDocument13 pagesCharacterization of Cell Wall Components of Wheat Bran Following Hydrothermal Pretreatment and FractionationAssefa DessieNo ratings yet
- Investigation of The Activity of Acinetobacter Calcoaceticus Biodemulsifier To Break Stable Water in Oil EmulsionsDocument7 pagesInvestigation of The Activity of Acinetobacter Calcoaceticus Biodemulsifier To Break Stable Water in Oil EmulsionsMohamed AlhayaniNo ratings yet
- Sustainability 15 06832Document20 pagesSustainability 15 06832Piero Omar Obregón EscuderoNo ratings yet
- C5RA17797ADocument29 pagesC5RA17797AABB HisseinNo ratings yet
- 1 s2.0 S025527012200143X MainDocument21 pages1 s2.0 S025527012200143X MainJHON ERIKNo ratings yet
- Thermophilic Biohydrogen Production From Optimized Enzymatic Pretreatment of Palm Oil Mill Effluent Via Box-Behnken DesignDocument8 pagesThermophilic Biohydrogen Production From Optimized Enzymatic Pretreatment of Palm Oil Mill Effluent Via Box-Behnken DesignosuolaleNo ratings yet
- Chapter 1Document6 pagesChapter 1likodblackbordNo ratings yet
- Valorization of Cassava Peels Waste Through Fermentation Process For Bioethanol Production: A ReviewDocument9 pagesValorization of Cassava Peels Waste Through Fermentation Process For Bioethanol Production: A ReviewIJAR JOURNALNo ratings yet
- Energies 16 00301Document13 pagesEnergies 16 00301DAVID ROSALESNo ratings yet
- Xing 2013Document8 pagesXing 2013Vijaya GosuNo ratings yet
- Effects of Dilute Acid and Alkaline Pretreatments On Enzymatic Saccharification of Palm Tree Trunk Waste For Bioethanol ProductionDocument10 pagesEffects of Dilute Acid and Alkaline Pretreatments On Enzymatic Saccharification of Palm Tree Trunk Waste For Bioethanol Productionsya viraNo ratings yet
- Processes 09 00436 v4Document23 pagesProcesses 09 00436 v4Maheesha GunathungaNo ratings yet
- Enhanced Mesophilic Anaerobic Digestion of PrimaryDocument21 pagesEnhanced Mesophilic Anaerobic Digestion of PrimaryVolute GroupNo ratings yet
- Membranes 13 00280Document15 pagesMembranes 13 00280rachel.amoilsNo ratings yet
- Using Dielectric Constant Measurement To Monitor Ethanol Fermentation and Anaerobic Co-Digestion of Lignocellulosic BiomassDocument20 pagesUsing Dielectric Constant Measurement To Monitor Ethanol Fermentation and Anaerobic Co-Digestion of Lignocellulosic Biomasszoltanjakoi.mkNo ratings yet
- 10.1007@s12649 019 00889 2Document11 pages10.1007@s12649 019 00889 2Jelian GraceNo ratings yet
- 1 s2.0 S0141813021018432 MainDocument10 pages1 s2.0 S0141813021018432 MainAnisa RatnasariNo ratings yet
- Anu 2020Document11 pagesAnu 2020Bandita DashNo ratings yet
- Maurya s13205-015-0279-4Document13 pagesMaurya s13205-015-0279-4Bandita DashNo ratings yet
- A Review On Palm Oil Mill Biogas Plant Wastewater Treatment Using Coagulation-OzonationDocument7 pagesA Review On Palm Oil Mill Biogas Plant Wastewater Treatment Using Coagulation-OzonationMaizan Sofia Binti Mohd MaarofNo ratings yet
- Recent Advances in Second Generation Bioethanol Production An Insight To Pretreatment, Saccharification and Fermentation ProcessesDocument11 pagesRecent Advances in Second Generation Bioethanol Production An Insight To Pretreatment, Saccharification and Fermentation ProcessesBryant CoolNo ratings yet
- Research ArticleDocument12 pagesResearch ArticlePranavNo ratings yet
- J Biortech 2017 12 033Document8 pagesJ Biortech 2017 12 033fajarNo ratings yet
- Valorization of Ibombax CeibDocument23 pagesValorization of Ibombax CeibAbdullah Nur HakamiNo ratings yet
- Influence of Sulfur Dioxide-Ethanol-Water Pretreatment On The Physicochemical Properties and Enzymatic Digestibility of Bamboo ResiduesDocument6 pagesInfluence of Sulfur Dioxide-Ethanol-Water Pretreatment On The Physicochemical Properties and Enzymatic Digestibility of Bamboo Residuesdesamek647No ratings yet
- Bioresource Technology: Sumit Kumar, Neerja Yadav, Lata Nain, Sunil Kumar KhareDocument4 pagesBioresource Technology: Sumit Kumar, Neerja Yadav, Lata Nain, Sunil Kumar KhareJosé BeltránNo ratings yet
- Oonkhanond 2017Document37 pagesOonkhanond 2017Gebby FebriliaNo ratings yet
- Process BiochemistryDocument4 pagesProcess BiochemistryIlham AkbarNo ratings yet
- Methyl CenlulozoDocument18 pagesMethyl CenlulozoMai QuỳnhNo ratings yet
- Putrino 2020Document7 pagesPutrino 2020Wayan ArnataNo ratings yet
- Energy Reports: Reem Shomal, Hiyam Hisham, Amal Mlhem, Rawan Hassan, Sulaiman Al-ZuhairDocument4 pagesEnergy Reports: Reem Shomal, Hiyam Hisham, Amal Mlhem, Rawan Hassan, Sulaiman Al-ZuhairHiyam HishamNo ratings yet
- F1C121039 - Finkyasti Nur Adman - Tugas Bahasa Inggris SainsDocument9 pagesF1C121039 - Finkyasti Nur Adman - Tugas Bahasa Inggris SainsFinkyasti Nur AdmanNo ratings yet
- Faculty of Applied SciencesDocument39 pagesFaculty of Applied SciencesashNo ratings yet
- Advances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsFrom EverandAdvances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsLalit Kumar SinghNo ratings yet
- John Ralph. 2010. Hydroxycinnamates in Lignification. Departments of Biochemistry and BiologicalDocument19 pagesJohn Ralph. 2010. Hydroxycinnamates in Lignification. Departments of Biochemistry and BiologicalAdhan AkbarNo ratings yet
- Fo Ohwoavworhua Dan Ta Adelkun. 2010. Non-Wood Fibre Production of Microcrystalline Cellulose From Sorghum CaudatumDocument7 pagesFo Ohwoavworhua Dan Ta Adelkun. 2010. Non-Wood Fibre Production of Microcrystalline Cellulose From Sorghum CaudatumAdhan AkbarNo ratings yet
- Mary L. Nelson, and Robert T. O'Connor. 1964. Relation of Certain Infrared Bands To Cellulose CrystallinityDocument17 pagesMary L. Nelson, and Robert T. O'Connor. 1964. Relation of Certain Infrared Bands To Cellulose CrystallinityAdhan AkbarNo ratings yet
- Muhaimin Muhaimin, Bayu Wiyantoko, Rahma Novia Putri, and Rika Rusitasari. 2018. Determination of Order ReactionDocument6 pagesMuhaimin Muhaimin, Bayu Wiyantoko, Rahma Novia Putri, and Rika Rusitasari. 2018. Determination of Order ReactionAdhan AkbarNo ratings yet
- Yulianti Sampora, Yenni Apriliany Devy, Dewi Sondari, Dan Athanasia Amanda Septevani. 2020. Simultaneous Pretreatment ProceDocument9 pagesYulianti Sampora, Yenni Apriliany Devy, Dewi Sondari, Dan Athanasia Amanda Septevani. 2020. Simultaneous Pretreatment ProceAdhan AkbarNo ratings yet
- Nattacha Paksung, Jens Pfersich, Pablo J. Arauzo, Dennis Jung, Dan Andrea Kruse. 2020. Structural EffectsDocument14 pagesNattacha Paksung, Jens Pfersich, Pablo J. Arauzo, Dennis Jung, Dan Andrea Kruse. 2020. Structural EffectsAdhan AkbarNo ratings yet
- Schwald, W. Dan Bobleter, O. Hydrothermolysis of Cellulose Under Static and Dynamic Conditions at High TemperaturesDocument15 pagesSchwald, W. Dan Bobleter, O. Hydrothermolysis of Cellulose Under Static and Dynamic Conditions at High TemperaturesAdhan AkbarNo ratings yet
- Ie8b00601 Si 001Document7 pagesIe8b00601 Si 001Adhan AkbarNo ratings yet
- Hazards and Risk Introduction To Pressure ProtectionDocument1 pageHazards and Risk Introduction To Pressure ProtectionAdhan AkbarNo ratings yet
- Extended Surface/ Finned Heat ExchangersDocument55 pagesExtended Surface/ Finned Heat ExchangersAdhan AkbarNo ratings yet
- Galihrinaldi, Linda Pulungan, Dono GuntoroDocument8 pagesGalihrinaldi, Linda Pulungan, Dono GuntoroAdhan AkbarNo ratings yet
- Evaluasi Kinerja Unit Crushing Plant Jaques Untuk Meningkatkan Produksi Granit Di Pt. Trimegah Perkasa Utama Kabupaten Karimun Kepulauan RiauDocument12 pagesEvaluasi Kinerja Unit Crushing Plant Jaques Untuk Meningkatkan Produksi Granit Di Pt. Trimegah Perkasa Utama Kabupaten Karimun Kepulauan RiauAdhan AkbarNo ratings yet
- Natural GasDocument13 pagesNatural GasAdhan AkbarNo ratings yet
- Rock and SoilDocument7 pagesRock and SoilEileen TeeNo ratings yet
- Catalog (Burner Nozzles) PDFDocument28 pagesCatalog (Burner Nozzles) PDFOCangaceiroNo ratings yet
- 2-Pager DuPont Delrin 511CPEDocument2 pages2-Pager DuPont Delrin 511CPEIrwan SaputraNo ratings yet
- 7 Types of Plastic Exist in Our Current Modern DaysDocument2 pages7 Types of Plastic Exist in Our Current Modern DaysTami AbordoNo ratings yet
- TN 36 Armature BandingDocument4 pagesTN 36 Armature Bandingpranav05No ratings yet
- PhysicsDocument189 pagesPhysicsSwetha INo ratings yet
- Stycast 1264 A/B: Low Viscosity, Transparent, Epoxy EncapsulantDocument2 pagesStycast 1264 A/B: Low Viscosity, Transparent, Epoxy EncapsulantRosli YaacobNo ratings yet
- Introduction To Pharmacology and Sources of DrugsDocument18 pagesIntroduction To Pharmacology and Sources of DrugskazeemNo ratings yet
- Thermal Cracking of Waste Engine Oil (Lubricant) at 430oc TemperatureDocument4 pagesThermal Cracking of Waste Engine Oil (Lubricant) at 430oc TemperatureAshok RathodNo ratings yet
- Product Development Flow Chart - Experimental Formulation DevelopmentDocument5 pagesProduct Development Flow Chart - Experimental Formulation DevelopmentNhư Ý TrịnhNo ratings yet
- Alchemist by Clockwork DragonDocument14 pagesAlchemist by Clockwork DragonTyler Longmuir0% (1)
- 2021 - WHO Key Technical Issues of Herbal Medicines With Reference To Interaction With Other Medicines With Others MedicinesDocument85 pages2021 - WHO Key Technical Issues of Herbal Medicines With Reference To Interaction With Other Medicines With Others MedicinesGabriela AlmeidaNo ratings yet
- 1 s2.0 S2352710223002152 MainDocument16 pages1 s2.0 S2352710223002152 MainEL AMAL MohamedNo ratings yet
- Tds Disperbyk-180 enDocument2 pagesTds Disperbyk-180 enSan HuỳnhNo ratings yet
- Glues PDFDocument11 pagesGlues PDFgalati12345No ratings yet
- Plancha Prensa AllianceDocument142 pagesPlancha Prensa AllianceHarrison JimenezNo ratings yet
- A Simplified Approach To Water-Hammer Analysis PDFDocument0 pagesA Simplified Approach To Water-Hammer Analysis PDFbtjajadi100% (1)
- Series 160S "S" Type Pitot Tubes: Operating InstructionsDocument2 pagesSeries 160S "S" Type Pitot Tubes: Operating InstructionsRazvanPopNo ratings yet
- TABLE 2 - Semi Essential AAsDocument37 pagesTABLE 2 - Semi Essential AAsHannalette GibanNo ratings yet
- Sharma 2012Document8 pagesSharma 2012yogi endiNo ratings yet
- The TITAN Grading Scheme For Used Containers: Grade ADocument3 pagesThe TITAN Grading Scheme For Used Containers: Grade ALe Cuong ThinhNo ratings yet
- CASE STUDY - Bhopal Gas TragedyDocument12 pagesCASE STUDY - Bhopal Gas TragedyAli Asif0% (1)
- Inconel 690Document8 pagesInconel 690Ronak shahNo ratings yet
- The Use of Gis in Assessing Exposure To Airborne Pollutants PDFDocument69 pagesThe Use of Gis in Assessing Exposure To Airborne Pollutants PDFstefania_iordache8578No ratings yet
- Structure, Properties and Uses of Industrially Important Oxides of BoronDocument18 pagesStructure, Properties and Uses of Industrially Important Oxides of BoronIshmita KaurNo ratings yet
- CHEM 430 NMR Spectroscopy Chapter 4Document95 pagesCHEM 430 NMR Spectroscopy Chapter 4Ravi GolNo ratings yet
- Glycerol Monooleate: Executive SummaryDocument16 pagesGlycerol Monooleate: Executive SummaryMarcelo Bacchetta100% (1)
- Magnetic Nanoparticles For Biomedical Applications: Yunsuk JoDocument69 pagesMagnetic Nanoparticles For Biomedical Applications: Yunsuk JoConstantin DenisaNo ratings yet
- Hydrolic Breaking SystemDocument56 pagesHydrolic Breaking SystemAnkush JainNo ratings yet
- Chemistry: Atoms, Molecules and IonsDocument60 pagesChemistry: Atoms, Molecules and Ionsjuliana100% (1)





































































































Download as pdf or txt
You might also like
- Alkaline Organosolv Pretreatment of Different Sorghum Stem Parts For Enhancing The Total Reducing Sugar Yields and P-Coumaric Acid ReleaseDocument14 pagesAlkaline Organosolv Pretreatment of Different Sorghum Stem Parts For Enhancing The Total Reducing Sugar Yields and P-Coumaric Acid Releaseuliseshbq7No ratings yet
- Industrially Relevant Hydrolyzability and Fermentability of Sugarcane Bagasse Improved Effectively by Glycerol Organosolv PretreatmentDocument13 pagesIndustrially Relevant Hydrolyzability and Fermentability of Sugarcane Bagasse Improved Effectively by Glycerol Organosolv PretreatmentHarshad Vinay SavantNo ratings yet
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- A Novel Recyclable Furoic Acid-Assisted Pretreatment For Sugarcane Bagasse Biorefinery in Co-Production of Xylooligosaccharides and GlucoseDocument8 pagesA Novel Recyclable Furoic Acid-Assisted Pretreatment For Sugarcane Bagasse Biorefinery in Co-Production of Xylooligosaccharides and GlucoseSthiden Sanabria ChaconNo ratings yet
- Feasibility Study of On-Site Solid-State Enzyme ProductionDocument15 pagesFeasibility Study of On-Site Solid-State Enzyme ProductionOuti ElinaNo ratings yet
- Influence of Sulfuric Acid Pretreatment and Inhibitor of Sugarcane Bagasse On The Production of Fermentable Sugar and EthanolDocument13 pagesInfluence of Sulfuric Acid Pretreatment and Inhibitor of Sugarcane Bagasse On The Production of Fermentable Sugar and EthanolJesusPJNo ratings yet
- Integrating Sugarcane Molasses Into Sequential Cellulosic Biofuel Production Based On SSF Process of High Solid LoadingDocument9 pagesIntegrating Sugarcane Molasses Into Sequential Cellulosic Biofuel Production Based On SSF Process of High Solid LoadingSamuel GamorNo ratings yet
- An Innovative Approach of Mixed Enzymatic Venture For 2G Ethanol Production From Lignocellulosic Feedstock PDFDocument10 pagesAn Innovative Approach of Mixed Enzymatic Venture For 2G Ethanol Production From Lignocellulosic Feedstock PDFRajiv Chandra RajakNo ratings yet
- International Journal of Biological Macromolecules Comparative Study 1-S2.0-S0141813019393699-MainDocument9 pagesInternational Journal of Biological Macromolecules Comparative Study 1-S2.0-S0141813019393699-MainOmar KHELILNo ratings yet
- Simultaneous Enzymatic Saccharification and Comminution For The Valorization of Lignocellulosic Biomass Toward Natural ProductsDocument11 pagesSimultaneous Enzymatic Saccharification and Comminution For The Valorization of Lignocellulosic Biomass Toward Natural ProductsSachin SharmaNo ratings yet
- Tongco2020 Article EnhancementOfHydrolysisAndBiogDocument9 pagesTongco2020 Article EnhancementOfHydrolysisAndBiogJim TsikasNo ratings yet
- Improvement of Enzymatic SaccharificationDocument11 pagesImprovement of Enzymatic SaccharificationpepandradeNo ratings yet
- International Journal of Biological MacromoleculesDocument7 pagesInternational Journal of Biological MacromoleculesEzanaNo ratings yet
- 14043-Article Text PDF-58576-2-10-20190902Document8 pages14043-Article Text PDF-58576-2-10-20190902R. VersteeghNo ratings yet
- 1 s2.0 S0360319922042458 MainDocument11 pages1 s2.0 S0360319922042458 Mainranjani093No ratings yet
- Isolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilDocument8 pagesIsolation and Screening of Hydrogen Producing Bacterial Strain From Sugarcane Bagasse Yard SoilATSNo ratings yet
- Enzymatic Digestibility of Pretreated Dewaxed Bamboo Residues As Feedstock For Bioethanol ProductionDocument7 pagesEnzymatic Digestibility of Pretreated Dewaxed Bamboo Residues As Feedstock For Bioethanol ProductionKevin Hernandez Duque100% (1)
- Materials: Multi-Functional Laccase Immobilized Hydrogel Microparticles For Efficient Removal of Bisphenol ADocument14 pagesMaterials: Multi-Functional Laccase Immobilized Hydrogel Microparticles For Efficient Removal of Bisphenol AcarolasbdNo ratings yet
- Accepted Manuscript: Bioresource TechnologyDocument32 pagesAccepted Manuscript: Bioresource TechnologyJulio MoralesNo ratings yet
- Structured Polyvinyl AlcoholZeoliteCarbon CompositDocument11 pagesStructured Polyvinyl AlcoholZeoliteCarbon CompositFarah TalibNo ratings yet
- Biochemical Characterization and SynergismDocument12 pagesBiochemical Characterization and SynergismJayesh PatidarNo ratings yet
- Biomass and Bioenergy: Research PaperDocument10 pagesBiomass and Bioenergy: Research PaperPaulina Lpz JrzNo ratings yet
- Sanjeetha Assignment 3 Proposal ReportDocument20 pagesSanjeetha Assignment 3 Proposal ReportSanjith KaurNo ratings yet
- 11403-Article Text PDF-34245-3-10-20180516Document12 pages11403-Article Text PDF-34245-3-10-20180516Mariana CampoNo ratings yet
- Dye RemovalDocument12 pagesDye RemovalaisyahNo ratings yet
- Research Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionDocument10 pagesResearch Article: Structural Analysis of Alkaline Pretreated Rice Straw For Ethanol ProductionShankar AcharNo ratings yet
- Characterization of Cell Wall Components of Wheat Bran Following Hydrothermal Pretreatment and FractionationDocument13 pagesCharacterization of Cell Wall Components of Wheat Bran Following Hydrothermal Pretreatment and FractionationAssefa DessieNo ratings yet
- Investigation of The Activity of Acinetobacter Calcoaceticus Biodemulsifier To Break Stable Water in Oil EmulsionsDocument7 pagesInvestigation of The Activity of Acinetobacter Calcoaceticus Biodemulsifier To Break Stable Water in Oil EmulsionsMohamed AlhayaniNo ratings yet
- Sustainability 15 06832Document20 pagesSustainability 15 06832Piero Omar Obregón EscuderoNo ratings yet
- C5RA17797ADocument29 pagesC5RA17797AABB HisseinNo ratings yet
- 1 s2.0 S025527012200143X MainDocument21 pages1 s2.0 S025527012200143X MainJHON ERIKNo ratings yet
- Thermophilic Biohydrogen Production From Optimized Enzymatic Pretreatment of Palm Oil Mill Effluent Via Box-Behnken DesignDocument8 pagesThermophilic Biohydrogen Production From Optimized Enzymatic Pretreatment of Palm Oil Mill Effluent Via Box-Behnken DesignosuolaleNo ratings yet
- Chapter 1Document6 pagesChapter 1likodblackbordNo ratings yet
- Valorization of Cassava Peels Waste Through Fermentation Process For Bioethanol Production: A ReviewDocument9 pagesValorization of Cassava Peels Waste Through Fermentation Process For Bioethanol Production: A ReviewIJAR JOURNALNo ratings yet
- Energies 16 00301Document13 pagesEnergies 16 00301DAVID ROSALESNo ratings yet
- Xing 2013Document8 pagesXing 2013Vijaya GosuNo ratings yet
- Effects of Dilute Acid and Alkaline Pretreatments On Enzymatic Saccharification of Palm Tree Trunk Waste For Bioethanol ProductionDocument10 pagesEffects of Dilute Acid and Alkaline Pretreatments On Enzymatic Saccharification of Palm Tree Trunk Waste For Bioethanol Productionsya viraNo ratings yet
- Processes 09 00436 v4Document23 pagesProcesses 09 00436 v4Maheesha GunathungaNo ratings yet
- Enhanced Mesophilic Anaerobic Digestion of PrimaryDocument21 pagesEnhanced Mesophilic Anaerobic Digestion of PrimaryVolute GroupNo ratings yet
- Membranes 13 00280Document15 pagesMembranes 13 00280rachel.amoilsNo ratings yet
- Using Dielectric Constant Measurement To Monitor Ethanol Fermentation and Anaerobic Co-Digestion of Lignocellulosic BiomassDocument20 pagesUsing Dielectric Constant Measurement To Monitor Ethanol Fermentation and Anaerobic Co-Digestion of Lignocellulosic Biomasszoltanjakoi.mkNo ratings yet
- 10.1007@s12649 019 00889 2Document11 pages10.1007@s12649 019 00889 2Jelian GraceNo ratings yet
- 1 s2.0 S0141813021018432 MainDocument10 pages1 s2.0 S0141813021018432 MainAnisa RatnasariNo ratings yet
- Anu 2020Document11 pagesAnu 2020Bandita DashNo ratings yet
- Maurya s13205-015-0279-4Document13 pagesMaurya s13205-015-0279-4Bandita DashNo ratings yet
- A Review On Palm Oil Mill Biogas Plant Wastewater Treatment Using Coagulation-OzonationDocument7 pagesA Review On Palm Oil Mill Biogas Plant Wastewater Treatment Using Coagulation-OzonationMaizan Sofia Binti Mohd MaarofNo ratings yet
- Recent Advances in Second Generation Bioethanol Production An Insight To Pretreatment, Saccharification and Fermentation ProcessesDocument11 pagesRecent Advances in Second Generation Bioethanol Production An Insight To Pretreatment, Saccharification and Fermentation ProcessesBryant CoolNo ratings yet
- Research ArticleDocument12 pagesResearch ArticlePranavNo ratings yet
- J Biortech 2017 12 033Document8 pagesJ Biortech 2017 12 033fajarNo ratings yet
- Valorization of Ibombax CeibDocument23 pagesValorization of Ibombax CeibAbdullah Nur HakamiNo ratings yet
- Influence of Sulfur Dioxide-Ethanol-Water Pretreatment On The Physicochemical Properties and Enzymatic Digestibility of Bamboo ResiduesDocument6 pagesInfluence of Sulfur Dioxide-Ethanol-Water Pretreatment On The Physicochemical Properties and Enzymatic Digestibility of Bamboo Residuesdesamek647No ratings yet
- Bioresource Technology: Sumit Kumar, Neerja Yadav, Lata Nain, Sunil Kumar KhareDocument4 pagesBioresource Technology: Sumit Kumar, Neerja Yadav, Lata Nain, Sunil Kumar KhareJosé BeltránNo ratings yet
- Oonkhanond 2017Document37 pagesOonkhanond 2017Gebby FebriliaNo ratings yet
- Process BiochemistryDocument4 pagesProcess BiochemistryIlham AkbarNo ratings yet
- Methyl CenlulozoDocument18 pagesMethyl CenlulozoMai QuỳnhNo ratings yet
- Putrino 2020Document7 pagesPutrino 2020Wayan ArnataNo ratings yet
- Energy Reports: Reem Shomal, Hiyam Hisham, Amal Mlhem, Rawan Hassan, Sulaiman Al-ZuhairDocument4 pagesEnergy Reports: Reem Shomal, Hiyam Hisham, Amal Mlhem, Rawan Hassan, Sulaiman Al-ZuhairHiyam HishamNo ratings yet
- F1C121039 - Finkyasti Nur Adman - Tugas Bahasa Inggris SainsDocument9 pagesF1C121039 - Finkyasti Nur Adman - Tugas Bahasa Inggris SainsFinkyasti Nur AdmanNo ratings yet
- Faculty of Applied SciencesDocument39 pagesFaculty of Applied SciencesashNo ratings yet
- Advances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsFrom EverandAdvances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsLalit Kumar SinghNo ratings yet
- John Ralph. 2010. Hydroxycinnamates in Lignification. Departments of Biochemistry and BiologicalDocument19 pagesJohn Ralph. 2010. Hydroxycinnamates in Lignification. Departments of Biochemistry and BiologicalAdhan AkbarNo ratings yet
- Fo Ohwoavworhua Dan Ta Adelkun. 2010. Non-Wood Fibre Production of Microcrystalline Cellulose From Sorghum CaudatumDocument7 pagesFo Ohwoavworhua Dan Ta Adelkun. 2010. Non-Wood Fibre Production of Microcrystalline Cellulose From Sorghum CaudatumAdhan AkbarNo ratings yet
- Mary L. Nelson, and Robert T. O'Connor. 1964. Relation of Certain Infrared Bands To Cellulose CrystallinityDocument17 pagesMary L. Nelson, and Robert T. O'Connor. 1964. Relation of Certain Infrared Bands To Cellulose CrystallinityAdhan AkbarNo ratings yet
- Muhaimin Muhaimin, Bayu Wiyantoko, Rahma Novia Putri, and Rika Rusitasari. 2018. Determination of Order ReactionDocument6 pagesMuhaimin Muhaimin, Bayu Wiyantoko, Rahma Novia Putri, and Rika Rusitasari. 2018. Determination of Order ReactionAdhan AkbarNo ratings yet
- Yulianti Sampora, Yenni Apriliany Devy, Dewi Sondari, Dan Athanasia Amanda Septevani. 2020. Simultaneous Pretreatment ProceDocument9 pagesYulianti Sampora, Yenni Apriliany Devy, Dewi Sondari, Dan Athanasia Amanda Septevani. 2020. Simultaneous Pretreatment ProceAdhan AkbarNo ratings yet
- Nattacha Paksung, Jens Pfersich, Pablo J. Arauzo, Dennis Jung, Dan Andrea Kruse. 2020. Structural EffectsDocument14 pagesNattacha Paksung, Jens Pfersich, Pablo J. Arauzo, Dennis Jung, Dan Andrea Kruse. 2020. Structural EffectsAdhan AkbarNo ratings yet
- Schwald, W. Dan Bobleter, O. Hydrothermolysis of Cellulose Under Static and Dynamic Conditions at High TemperaturesDocument15 pagesSchwald, W. Dan Bobleter, O. Hydrothermolysis of Cellulose Under Static and Dynamic Conditions at High TemperaturesAdhan AkbarNo ratings yet
- Ie8b00601 Si 001Document7 pagesIe8b00601 Si 001Adhan AkbarNo ratings yet
- Hazards and Risk Introduction To Pressure ProtectionDocument1 pageHazards and Risk Introduction To Pressure ProtectionAdhan AkbarNo ratings yet
- Extended Surface/ Finned Heat ExchangersDocument55 pagesExtended Surface/ Finned Heat ExchangersAdhan AkbarNo ratings yet
- Galihrinaldi, Linda Pulungan, Dono GuntoroDocument8 pagesGalihrinaldi, Linda Pulungan, Dono GuntoroAdhan AkbarNo ratings yet
- Evaluasi Kinerja Unit Crushing Plant Jaques Untuk Meningkatkan Produksi Granit Di Pt. Trimegah Perkasa Utama Kabupaten Karimun Kepulauan RiauDocument12 pagesEvaluasi Kinerja Unit Crushing Plant Jaques Untuk Meningkatkan Produksi Granit Di Pt. Trimegah Perkasa Utama Kabupaten Karimun Kepulauan RiauAdhan AkbarNo ratings yet
- Natural GasDocument13 pagesNatural GasAdhan AkbarNo ratings yet
- Rock and SoilDocument7 pagesRock and SoilEileen TeeNo ratings yet
- Catalog (Burner Nozzles) PDFDocument28 pagesCatalog (Burner Nozzles) PDFOCangaceiroNo ratings yet
- 2-Pager DuPont Delrin 511CPEDocument2 pages2-Pager DuPont Delrin 511CPEIrwan SaputraNo ratings yet
- 7 Types of Plastic Exist in Our Current Modern DaysDocument2 pages7 Types of Plastic Exist in Our Current Modern DaysTami AbordoNo ratings yet
- TN 36 Armature BandingDocument4 pagesTN 36 Armature Bandingpranav05No ratings yet
- PhysicsDocument189 pagesPhysicsSwetha INo ratings yet
- Stycast 1264 A/B: Low Viscosity, Transparent, Epoxy EncapsulantDocument2 pagesStycast 1264 A/B: Low Viscosity, Transparent, Epoxy EncapsulantRosli YaacobNo ratings yet
- Introduction To Pharmacology and Sources of DrugsDocument18 pagesIntroduction To Pharmacology and Sources of DrugskazeemNo ratings yet
- Thermal Cracking of Waste Engine Oil (Lubricant) at 430oc TemperatureDocument4 pagesThermal Cracking of Waste Engine Oil (Lubricant) at 430oc TemperatureAshok RathodNo ratings yet
- Product Development Flow Chart - Experimental Formulation DevelopmentDocument5 pagesProduct Development Flow Chart - Experimental Formulation DevelopmentNhư Ý TrịnhNo ratings yet
- Alchemist by Clockwork DragonDocument14 pagesAlchemist by Clockwork DragonTyler Longmuir0% (1)
- 2021 - WHO Key Technical Issues of Herbal Medicines With Reference To Interaction With Other Medicines With Others MedicinesDocument85 pages2021 - WHO Key Technical Issues of Herbal Medicines With Reference To Interaction With Other Medicines With Others MedicinesGabriela AlmeidaNo ratings yet
- 1 s2.0 S2352710223002152 MainDocument16 pages1 s2.0 S2352710223002152 MainEL AMAL MohamedNo ratings yet
- Tds Disperbyk-180 enDocument2 pagesTds Disperbyk-180 enSan HuỳnhNo ratings yet
- Glues PDFDocument11 pagesGlues PDFgalati12345No ratings yet
- Plancha Prensa AllianceDocument142 pagesPlancha Prensa AllianceHarrison JimenezNo ratings yet
- A Simplified Approach To Water-Hammer Analysis PDFDocument0 pagesA Simplified Approach To Water-Hammer Analysis PDFbtjajadi100% (1)
- Series 160S "S" Type Pitot Tubes: Operating InstructionsDocument2 pagesSeries 160S "S" Type Pitot Tubes: Operating InstructionsRazvanPopNo ratings yet
- TABLE 2 - Semi Essential AAsDocument37 pagesTABLE 2 - Semi Essential AAsHannalette GibanNo ratings yet
- Sharma 2012Document8 pagesSharma 2012yogi endiNo ratings yet
- The TITAN Grading Scheme For Used Containers: Grade ADocument3 pagesThe TITAN Grading Scheme For Used Containers: Grade ALe Cuong ThinhNo ratings yet
- CASE STUDY - Bhopal Gas TragedyDocument12 pagesCASE STUDY - Bhopal Gas TragedyAli Asif0% (1)
- Inconel 690Document8 pagesInconel 690Ronak shahNo ratings yet
- The Use of Gis in Assessing Exposure To Airborne Pollutants PDFDocument69 pagesThe Use of Gis in Assessing Exposure To Airborne Pollutants PDFstefania_iordache8578No ratings yet
- Structure, Properties and Uses of Industrially Important Oxides of BoronDocument18 pagesStructure, Properties and Uses of Industrially Important Oxides of BoronIshmita KaurNo ratings yet
- CHEM 430 NMR Spectroscopy Chapter 4Document95 pagesCHEM 430 NMR Spectroscopy Chapter 4Ravi GolNo ratings yet
- Glycerol Monooleate: Executive SummaryDocument16 pagesGlycerol Monooleate: Executive SummaryMarcelo Bacchetta100% (1)
- Magnetic Nanoparticles For Biomedical Applications: Yunsuk JoDocument69 pagesMagnetic Nanoparticles For Biomedical Applications: Yunsuk JoConstantin DenisaNo ratings yet
- Hydrolic Breaking SystemDocument56 pagesHydrolic Breaking SystemAnkush JainNo ratings yet
- Chemistry: Atoms, Molecules and IonsDocument60 pagesChemistry: Atoms, Molecules and Ionsjuliana100% (1)