Download as pdf or txt
You might also like
- Robert Plutchik (Auth.), Manfred Clynes, Jaak Panksepp (Eds.) - Emotions and Psychopathology-Springer US (1988)Document336 pagesRobert Plutchik (Auth.), Manfred Clynes, Jaak Panksepp (Eds.) - Emotions and Psychopathology-Springer US (1988)Lalón Gitud De Onda100% (1)
- Dulce BattleDocument98 pagesDulce BattleMarc St-Germain100% (3)
- 6 Enzymes and Cellular Regulation-SDocument5 pages6 Enzymes and Cellular Regulation-Sapi-502781581No ratings yet
- Animal Habitat Diorama Project InstructionsDocument7 pagesAnimal Habitat Diorama Project Instructionsapi-293342816No ratings yet
- Cells 12 00321Document30 pagesCells 12 00321uttamNo ratings yet
- GeneDocument10 pagesGeneLýDiệuNo ratings yet
- JHep 2010Document10 pagesJHep 2010wangyu81No ratings yet
- Hematopoietic Stem and ProgenitorCellsDocument10 pagesHematopoietic Stem and ProgenitorCellsRuxandra MesarosNo ratings yet
- POT0024 - 1117B - StemCell Insider Issue 8 (Expansion)Document2 pagesPOT0024 - 1117B - StemCell Insider Issue 8 (Expansion)Francisco MoralesNo ratings yet
- 42 Uniform Scale Independent Gene Transfer To Striated Muscle After Transvenular Extravasation of VectorDocument2 pages42 Uniform Scale Independent Gene Transfer To Striated Muscle After Transvenular Extravasation of VectorMuinimula aluminiuM (MuminiminimuM)No ratings yet
- Neural Differentiation of Human Pluripotent Stem CellsDocument12 pagesNeural Differentiation of Human Pluripotent Stem Cellsilmiah neurologiNo ratings yet
- Silibinin Inhibits Proliferation and Migration of Human Hepatic Stellate LX-2 CellsDocument3 pagesSilibinin Inhibits Proliferation and Migration of Human Hepatic Stellate LX-2 CellsSILVIA BAQUEIRONo ratings yet
- (Review) (Biological Chemistry) Bernd Giebel Et Michael Punzel 2008Document12 pages(Review) (Biological Chemistry) Bernd Giebel Et Michael Punzel 2008Priscilla FreschuNo ratings yet
- Wharton's JellyDocument13 pagesWharton's JellyZ MNo ratings yet
- Ijsc 13 001 2Document12 pagesIjsc 13 001 2dimiz77No ratings yet
- SCT3 6 0864Document13 pagesSCT3 6 0864Z MNo ratings yet
- Zeng 2013Document7 pagesZeng 2013angela_karenina_1No ratings yet
- Cancer 1Document8 pagesCancer 1JoseRodrigoSuarezCamposNo ratings yet
- LCH TargetDocument5 pagesLCH TargetSeham GoharNo ratings yet
- Mehta 2011Document10 pagesMehta 2011Giulia AndreeaNo ratings yet
- NIH Public AccessDocument18 pagesNIH Public AccessSofia BottiNo ratings yet
- The Physical Microenvironment of Hematopoietic Stem Cells and Its Emerging Roles in Engineering ApplicationsDocument13 pagesThe Physical Microenvironment of Hematopoietic Stem Cells and Its Emerging Roles in Engineering ApplicationsDemie PearlNo ratings yet
- Gene TherapyDocument6 pagesGene TherapyHafizah RamliNo ratings yet
- Hematopoietic Stem Cells: Gordon KellerDocument7 pagesHematopoietic Stem Cells: Gordon KellerPisi MicaNo ratings yet
- BBA - Molecular Cell ResearchDocument11 pagesBBA - Molecular Cell ResearchPUSKESMAS KECAMATAN KEPULAUAN SERIBU UTARANo ratings yet
- BPM EN Produccion de FibroblastosDocument14 pagesBPM EN Produccion de FibroblastosAndrea Garcia CaycedoNo ratings yet
- Artigo Biotecnologia Tecido AdiposoDocument15 pagesArtigo Biotecnologia Tecido AdiposoLuiz Gabriel Ferreira da SilvaNo ratings yet
- Changes in The Cell Surface Markers During NormalDocument10 pagesChanges in The Cell Surface Markers During NormalMaw BerryNo ratings yet
- Dimensions of Neutrophil Life and Fate 2020Document7 pagesDimensions of Neutrophil Life and Fate 2020brunareis1999No ratings yet
- Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and ApplicationsDocument30 pagesAdvances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and ApplicationsnembutalNo ratings yet
- Cells 12 02765 v2Document22 pagesCells 12 02765 v2Naga MuthuNo ratings yet
- Cell Regeneration 2012Document10 pagesCell Regeneration 2012ИгорьNo ratings yet
- Science Ade6967Document9 pagesScience Ade69670728lrhNo ratings yet
- Base Editing of Hematopoietic Stem Cells Rescues Sickle Cell Disease in MiceDocument42 pagesBase Editing of Hematopoietic Stem Cells Rescues Sickle Cell Disease in MiceOsvaldo VillarNo ratings yet
- Chapter 3Document39 pagesChapter 3hedieh abedNo ratings yet
- Jurnal Metastasis Paru 2Document25 pagesJurnal Metastasis Paru 2widiastrikNo ratings yet
- IJKD - News Letter1 20 26Document8 pagesIJKD - News Letter1 20 26Kapil SejpalNo ratings yet
- Pnas 1911951117Document11 pagesPnas 1911951117Lucero RamirezNo ratings yet
- Increased Mab Production in Amplified CHO Cell Lines Is Associated With Increased Interaction of CREB1 With Transgene PromoterDocument9 pagesIncreased Mab Production in Amplified CHO Cell Lines Is Associated With Increased Interaction of CREB1 With Transgene PromoterSusan SharfsteinNo ratings yet
- S74 Poster Abstracts: Table 1Document1 pageS74 Poster Abstracts: Table 1HanaNo ratings yet
- 2023 Article 529Document22 pages2023 Article 529amirNo ratings yet
- Cell Reprogramming: Expectations and Challenges For Chemistry in Stem Cell Biology and Regenerative MedicineDocument8 pagesCell Reprogramming: Expectations and Challenges For Chemistry in Stem Cell Biology and Regenerative MedicinenembutalNo ratings yet
- Fimmu 13 814030Document15 pagesFimmu 13 814030yunitaknNo ratings yet
- Gene TherapyDocument10 pagesGene TherapyIsabella MejiaNo ratings yet
- Hepatitis B Research PapersDocument4 pagesHepatitis B Research Papersafnhgewvmftbsm100% (1)
- ACTN4-자궁경부암 관계 입증 논문-3Document12 pagesACTN4-자궁경부암 관계 입증 논문-3김승훈(이과대학 화학)No ratings yet
- NatureDocument32 pagesNaturezhouqunli514No ratings yet
- CA A Cancer J Clinicians - March April 1988 - Virji - Tumor Markers in Cancer Diagnosis and PrognosisDocument23 pagesCA A Cancer J Clinicians - March April 1988 - Virji - Tumor Markers in Cancer Diagnosis and PrognosisIssahaku AbdallahNo ratings yet
- The Concept of Radiation-Enhanced Stem Cell Differentiation: Research ArticleDocument8 pagesThe Concept of Radiation-Enhanced Stem Cell Differentiation: Research ArticledwinugrohojuandaNo ratings yet
- A Recombinant Human Anti-Platelet SCFV Antibody Produced in Pichia Pastoris For Atheroma TargetingDocument18 pagesA Recombinant Human Anti-Platelet SCFV Antibody Produced in Pichia Pastoris For Atheroma TargetingSyahputraWibowoNo ratings yet
- Yu Et Al-2018-Journal of Cellular and Molecular MedicineDocument9 pagesYu Et Al-2018-Journal of Cellular and Molecular MedicineHanzZzAravlaNo ratings yet
- Nihms 1807736Document15 pagesNihms 1807736sirfengchenNo ratings yet
- Identification of Cancer-Related Mutations in Human Pluripotent Stem Cells Using RNA-seq AnalysisDocument16 pagesIdentification of Cancer-Related Mutations in Human Pluripotent Stem Cells Using RNA-seq AnalysisgangligonNo ratings yet
- 2020 The Application of Cell Surface Markers To Demarcate Distinct Human Pluripotent States (Experimental Cell Research)Document9 pages2020 The Application of Cell Surface Markers To Demarcate Distinct Human Pluripotent States (Experimental Cell Research)Thakoon TtsNo ratings yet
- Embedding The Future of Regenerative Medicine Into The Open Epigenomic Landscape of Pluripotent Human Embryonic Stem CellsDocument29 pagesEmbedding The Future of Regenerative Medicine Into The Open Epigenomic Landscape of Pluripotent Human Embryonic Stem CellsMili GiménezNo ratings yet
- Clonally Resolved Single Cell Multi Omics Identifies Routes of 2023 Cell STDocument25 pagesClonally Resolved Single Cell Multi Omics Identifies Routes of 2023 Cell STdimiz77No ratings yet
- Recombinant Human G-CSF Production As A Protein Based Drug Candidate For Hematology and OncologyDocument9 pagesRecombinant Human G-CSF Production As A Protein Based Drug Candidate For Hematology and OncologyZminasariNo ratings yet
- HomingDocument14 pagesHomingMaría P SNo ratings yet
- Poster Presentations Experimental Hematology 44 (2016) S56-S110Document1 pagePoster Presentations Experimental Hematology 44 (2016) S56-S110aymenNo ratings yet
- Stem Cell: The Spleen Contributes Stem Cells To Peripheral Blood Stem Cell TransplantsDocument4 pagesStem Cell: The Spleen Contributes Stem Cells To Peripheral Blood Stem Cell TransplantsAnuj MittalNo ratings yet
- Multi Lineage Progenitor Cells Ped 1660 06Document2 pagesMulti Lineage Progenitor Cells Ped 1660 06608943No ratings yet
- IUBMB Life - 2020 - Pozzi - Cancer Stem Cell Enrichment Is Associated With Enhancement of Nicotinamide N MethyltransferaseDocument11 pagesIUBMB Life - 2020 - Pozzi - Cancer Stem Cell Enrichment Is Associated With Enhancement of Nicotinamide N MethyltransferasePatricia SolarNo ratings yet
- Eshtiaghi 2019Document6 pagesEshtiaghi 2019dimiz77No ratings yet
- Diagnosis of Blood and Bone Marrow DisordersFrom EverandDiagnosis of Blood and Bone Marrow DisordersSa A. WangNo ratings yet
- Thesis Statement For HoneybeesDocument5 pagesThesis Statement For Honeybeesgjgcnp6z100% (2)
- Navarro College Medical Laboratory Technology Degree PlanDocument1 pageNavarro College Medical Laboratory Technology Degree PlanNavarro CollegeNo ratings yet
- Glycolysis Krebscycle Practice Questions SCDocument2 pagesGlycolysis Krebscycle Practice Questions SCapi-323720899No ratings yet
- Probni Test 1. Godina - Ina KlipaDocument4 pagesProbni Test 1. Godina - Ina KlipaMickoNo ratings yet
- Myo 5 ADocument26 pagesMyo 5 Aapi-221640706No ratings yet
- Chromatography REVISED2016Document10 pagesChromatography REVISED2016Abby LumanglasNo ratings yet
- Checklist - Bathing An Adult ClientDocument1 pageChecklist - Bathing An Adult ClientDonaJeanNo ratings yet
- Summary of Pub Med Articles Protandim PDFDocument2 pagesSummary of Pub Med Articles Protandim PDFLeslee MasonNo ratings yet
- Experiment 3 An O-Toluidine Method For Blood Glucose DeterminationDocument9 pagesExperiment 3 An O-Toluidine Method For Blood Glucose DeterminationAlmas MalikNo ratings yet
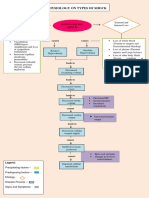
- Pathophysiology On Types of ShockDocument4 pagesPathophysiology On Types of ShockJessa Mae Alforque AsentistaNo ratings yet
- Anatomy and Physiology CNS Brain and Spinal Cord Chart Completed AP BiologyDocument12 pagesAnatomy and Physiology CNS Brain and Spinal Cord Chart Completed AP BiologyTiffany GallinaNo ratings yet
- Introduction To MorphologyDocument17 pagesIntroduction To MorphologydibabasarNo ratings yet
- Ankylosing SpondylitisDocument19 pagesAnkylosing SpondylitisZulhida YuniNo ratings yet
- Attention and Concentration in SportsDocument30 pagesAttention and Concentration in SportsArundhathy GopakumarNo ratings yet
- Radtke-Schuller2016 Article BrainAtlasOfTheMongolianGerbilDocument272 pagesRadtke-Schuller2016 Article BrainAtlasOfTheMongolianGerbilCharlotte NordenNo ratings yet
- P (3) Centrifuge Uses and MaitenanceDocument17 pagesP (3) Centrifuge Uses and MaitenanceSaadNo ratings yet
- Diffenrece Between Filipino TraditionalDocument19 pagesDiffenrece Between Filipino TraditionalJansen ValdezNo ratings yet
- New Case Study Booklet-1Document13 pagesNew Case Study Booklet-1Dr Sumant SharmaNo ratings yet
- Hipotiroidisme Pasca Radioterapi Untuk Karsinoma NasofaringDocument28 pagesHipotiroidisme Pasca Radioterapi Untuk Karsinoma NasofaringDeandles WattimuryNo ratings yet
- PhyloCode v5 BetaDocument69 pagesPhyloCode v5 Betagobxs4b1iNo ratings yet
- Life Sciences Part 2 CSIR JRF NET GATE DBT PDFDocument167 pagesLife Sciences Part 2 CSIR JRF NET GATE DBT PDFsaranya100% (2)
- FCI Previous Question PapersDocument5 pagesFCI Previous Question PapersRohit BeriNo ratings yet
- Rheedea19 73-74Document2 pagesRheedea19 73-74manudevkmadhavanNo ratings yet
- Core Program Spring 2022 23 Course Details Jan09Document54 pagesCore Program Spring 2022 23 Course Details Jan09movefe8960No ratings yet
- Hia HraDocument30 pagesHia Hrashintia febrianiNo ratings yet
- Diamond 1997Document457 pagesDiamond 1997Learning Club100% (3)