Download as pdf or txt
You might also like
- Dwnload Full Microbiology A Laboratory Manual 11th Edition Cappuccino Solutions Manual PDFDocument36 pagesDwnload Full Microbiology A Laboratory Manual 11th Edition Cappuccino Solutions Manual PDFyoreoakling.9vtk0y100% (13)
- Pollination Class 12 Biology ProjectDocument18 pagesPollination Class 12 Biology ProjectSiddhartha Gautama76% (89)
- Qdoc - Tips Pollination Class 12 Biology ProjectDocument18 pagesQdoc - Tips Pollination Class 12 Biology ProjectPranav SNo ratings yet
- Why Animals Don't Get Heart Attacks - But - Rath, Matthias, M.D PDFDocument324 pagesWhy Animals Don't Get Heart Attacks - But - Rath, Matthias, M.D PDFAnonymous Jap77xvqPK100% (3)
- Experiment 1 Pollen GerminationDocument2 pagesExperiment 1 Pollen Germinationshr sem100% (3)
- Mechanisms of Development: ReviewDocument5 pagesMechanisms of Development: ReviewHelena Ribeiro SouzaNo ratings yet
- PollinationDocument22 pagesPollinationNAVEEN DHIMAN100% (1)
- Department of Biology: Jawahar Navodaya Vidyalaya ShegaonDocument25 pagesDepartment of Biology: Jawahar Navodaya Vidyalaya Shegaonprasad panjabrao mapariNo ratings yet
- Pollination Class 12 Biology ProjectDocument18 pagesPollination Class 12 Biology ProjectRajat Rawat0% (1)
- Adobe Scan Oct 31, 2023Document25 pagesAdobe Scan Oct 31, 2023amanazim404No ratings yet
- KvmzupollinationprojectDocument18 pagesKvmzupollinationprojectK. K KumarNo ratings yet
- ISSN 1679-9216 (Online)Document19 pagesISSN 1679-9216 (Online)Rachel HayonNo ratings yet
- Biology Investigatory ProjectDocument24 pagesBiology Investigatory ProjectAhalya Bai SahooNo ratings yet
- PalynologyDocument18 pagesPalynologyDr. Prashant SinghNo ratings yet
- Journal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacDocument14 pagesJournal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacMashai LesenyehoNo ratings yet
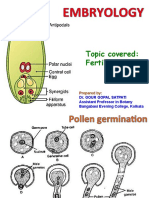
- Topic Covered: Fertilization: Core Course 6 Semester IiiDocument9 pagesTopic Covered: Fertilization: Core Course 6 Semester Iiisiddharth pandeyNo ratings yet
- Ilide - Info Pollination Class 12 Biology Project PRDocument18 pagesIlide - Info Pollination Class 12 Biology Project PRThuji ZangpoNo ratings yet
- PreciadoGarcía Fátima BiologyII 45BBDocument5 pagesPreciadoGarcía Fátima BiologyII 45BBFatima PreciadoNo ratings yet
- Lab Rep 13Document17 pagesLab Rep 13Reign RosadaNo ratings yet
- Double FertilizationDocument4 pagesDouble FertilizationHemanta JenaNo ratings yet
- Pollination Class 12 Biology ProjectDocument21 pagesPollination Class 12 Biology ProjectASHOK SAGAR85% (20)
- Tomlinson 1992. Podocarpaceae ConeDocument18 pagesTomlinson 1992. Podocarpaceae ConejorbelocoNo ratings yet
- A Comparative Cytological Analysis of Fu20160623 13956 q2w8t5 With Cover PageDocument15 pagesA Comparative Cytological Analysis of Fu20160623 13956 q2w8t5 With Cover PageCristián MaximilianoNo ratings yet
- Pollination in Sunflower and LegumDocument18 pagesPollination in Sunflower and Legumkanha kumarNo ratings yet
- DiabetesDocument19 pagesDiabetesR.KABILANNo ratings yet
- Pathogens 10 01029Document10 pagesPathogens 10 01029Vicky PokemonNo ratings yet
- Double Fertilization - WikipediaDocument27 pagesDouble Fertilization - WikipediaNaniNo ratings yet
- PBT NotesDocument5 pagesPBT NotesYogish HegdeNo ratings yet
- Double Fertilization - WikipediaDocument27 pagesDouble Fertilization - WikipediaBashiir NuurNo ratings yet
- Dvdy24087 5 Final As PublishedDocument12 pagesDvdy24087 5 Final As PublishedTahseen FatimaNo ratings yet
- Pollen Germination On Stigma Through A Permanent SlideDocument1 pagePollen Germination On Stigma Through A Permanent SlideGolu YadavNo ratings yet
- Reproduction in OrganismsDocument15 pagesReproduction in OrganismsAlakesh Coldplay KalitaNo ratings yet
- Pollen-Pistil Interaction: Pollen Attachment and HydrationDocument6 pagesPollen-Pistil Interaction: Pollen Attachment and HydrationSreeja Raj100% (1)
- Easily Automate Your Pipe Ing: Connected Smart PipettingDocument10 pagesEasily Automate Your Pipe Ing: Connected Smart PipettingSmruti ParhiNo ratings yet
- Biology Investigatory ProjectDocument19 pagesBiology Investigatory ProjectPriyanks Rout50% (2)
- Dallai Mating Zorotypus2013Document15 pagesDallai Mating Zorotypus2013cr rNo ratings yet
- Seijjjj 378777y36262626363. 4444 22373737374 A - U8ujb Vjhdjdjd. Bbwqwqwoo Rchins - As - Model - System - For - Studying - DevelopmentDocument7 pagesSeijjjj 378777y36262626363. 4444 22373737374 A - U8ujb Vjhdjdjd. Bbwqwqwoo Rchins - As - Model - System - For - Studying - DevelopmentRed DiggerNo ratings yet
- The Avian OrganizerDocument7 pagesThe Avian OrganizerTamara OliveiraNo ratings yet
- Lepidoptera Genetics: International Series of Monographs in Pure and Applied Biology: ZoologyFrom EverandLepidoptera Genetics: International Series of Monographs in Pure and Applied Biology: ZoologyRating: 5 out of 5 stars5/5 (1)
- Pollination WikipediaDocument12 pagesPollination WikipediaAnirban MallickNo ratings yet
- Green Beginnings - Pattern Formation in The Early Plant EmbryoDocument27 pagesGreen Beginnings - Pattern Formation in The Early Plant EmbryoMark DenhamNo ratings yet
- Pollination and FertilizationDocument12 pagesPollination and Fertilizationmiraenriquez78No ratings yet
- Unit 11Document33 pagesUnit 11shiva kumar goudNo ratings yet
- InokulasiDocument15 pagesInokulasiMhd WldNo ratings yet
- 2021 ManualDocument32 pages2021 ManualBalaji G.JNo ratings yet
- Microbiology A Laboratory Manual 11th Edition Cappuccino Solutions Manual 1Document36 pagesMicrobiology A Laboratory Manual 11th Edition Cappuccino Solutions Manual 1joshuawongawzspdkxjb100% (29)
- Pollen-Pistil Interaction: Recognition of Compatible PollenDocument3 pagesPollen-Pistil Interaction: Recognition of Compatible PollenHarpreetNo ratings yet
- Phase and Caste Determination in Insects: Endocrine AspectsFrom EverandPhase and Caste Determination in Insects: Endocrine AspectsMartin LüscherNo ratings yet
- Biology Investigatory Project: PollinationDocument30 pagesBiology Investigatory Project: PollinationMohamed LaqinNo ratings yet
- Early Land Plants 2019Document17 pagesEarly Land Plants 2019Joseane FerrazNo ratings yet
- Final BiologyDocument3 pagesFinal BiologySunidhi SinghNo ratings yet
- Hyphal and Mycelial ConsciousnessDocument3 pagesHyphal and Mycelial Consciousness202200024No ratings yet
- Prepare The Materials Wait For 2 Months Observe The Modified Stems, Leaves and RootsDocument8 pagesPrepare The Materials Wait For 2 Months Observe The Modified Stems, Leaves and RootsBanana LatteNo ratings yet
- Bot01 Lec4Document11 pagesBot01 Lec4Jan Leo QuinioNo ratings yet
- EvoDevo Milestones 1-12Document24 pagesEvoDevo Milestones 1-12Giovana Brito De LimaNo ratings yet
- Microbiology A Laboratory Manual 11th Edition Cappuccino Solutions ManualDocument36 pagesMicrobiology A Laboratory Manual 11th Edition Cappuccino Solutions Manualbegravebasnetvp8su100% (23)
- The Veterinary Journal: V. Hall, K. Hinrichs, G. Lazzari, D.H. Betts, P. HyttelDocument15 pagesThe Veterinary Journal: V. Hall, K. Hinrichs, G. Lazzari, D.H. Betts, P. HyttelJuniClaudia13No ratings yet
- Articulo en Ingles D. MelanogasterDocument6 pagesArticulo en Ingles D. Melanogasterzafiro TorresNo ratings yet
- 1 s2.0 S0093691X13003865 MainDocument7 pages1 s2.0 S0093691X13003865 Maincarrerajm22No ratings yet
- App Phal 2010Document8 pagesApp Phal 2010skullabyNo ratings yet
- Parasita No Intestino Do MosquitoDocument8 pagesParasita No Intestino Do MosquitoGustavo FelpeNo ratings yet
- Youarewhatyoueat MadisonmacdonaldDocument6 pagesYouarewhatyoueat Madisonmacdonaldapi-305117392No ratings yet
- Timeline ActivityDocument2 pagesTimeline ActivityhanniemaelimonNo ratings yet
- Mitosis Through Microscope Science 2003Document7 pagesMitosis Through Microscope Science 2003Fran IgarzabalNo ratings yet
- 2014 Morin PantoeaDocument5 pages2014 Morin PantoeaArnab PurohitNo ratings yet
- Problem Set 2 EndocrinologyDocument12 pagesProblem Set 2 EndocrinologyJessica EtieneNo ratings yet
- 1 Study Guide - Unit 2Document3 pages1 Study Guide - Unit 2slimjimonfire5682No ratings yet
- High-Resolution Screening of Metabolite-Like Lead LibrariesDocument114 pagesHigh-Resolution Screening of Metabolite-Like Lead LibrariesBuscador AlfaNo ratings yet
- The Antioxidant, Immunomodulatory, and Anti Inflammatory Activities of Spirulina: An OverviewDocument24 pagesThe Antioxidant, Immunomodulatory, and Anti Inflammatory Activities of Spirulina: An OverviewPlacido Rojas FrancoNo ratings yet
- Class 10 CH 8 Important QuestionsDocument31 pagesClass 10 CH 8 Important QuestionsagrimNo ratings yet
- Forensic Video WorksheetDocument2 pagesForensic Video Worksheetshadow12345No ratings yet
- Artepillin C As An Outstanding Phenolic Compound of Brazilian Green Propolis For Disease Treatment: A Review On Pharmacological AspectsDocument13 pagesArtepillin C As An Outstanding Phenolic Compound of Brazilian Green Propolis For Disease Treatment: A Review On Pharmacological AspectslyviaNo ratings yet
- A Laboratory Manual of Invertebrate Zoology 1920Document248 pagesA Laboratory Manual of Invertebrate Zoology 1920PramestiDwiPutri100% (1)
- Lupus Nephritis Diagnosis and ClassificationDocument37 pagesLupus Nephritis Diagnosis and ClassificationElena StepanNo ratings yet
- Diagnosis of Canine Claw Disease - A Prospective Study of 24 DogsDocument9 pagesDiagnosis of Canine Claw Disease - A Prospective Study of 24 DogsjenNo ratings yet
- ASSESSMENT TASK 1-CHCCCS041-CHC33021-CYCLE A-RTO WORKS-V1.0 2023 (Review)Document38 pagesASSESSMENT TASK 1-CHCCCS041-CHC33021-CYCLE A-RTO WORKS-V1.0 2023 (Review)mutnguyen141No ratings yet
- Novartis - Case StudyDocument5 pagesNovartis - Case Studydhrubo_tara25No ratings yet
- Taligaman National High SchoolDocument3 pagesTaligaman National High SchoolMelvin CabonegroNo ratings yet
- Bahasa Inggris XiiDocument6 pagesBahasa Inggris Xiinurmala sariNo ratings yet
- Thesis PCRDocument8 pagesThesis PCRgjfs5mtv100% (2)
- Module 1 - Introduction To BiologyDocument7 pagesModule 1 - Introduction To BiologyYsabela Dela PazNo ratings yet
- Michael Banton The Vertical and Horizontal Dimensions of The Word RaceDocument14 pagesMichael Banton The Vertical and Horizontal Dimensions of The Word RaceAldair RodriguesNo ratings yet
- Maths Maths Maths Maths: VIDAAT (2023-2024) Proposed SyllabusDocument6 pagesMaths Maths Maths Maths: VIDAAT (2023-2024) Proposed SyllabusOneShotNo ratings yet
- Pathogenic Gram Positive BacteriaDocument34 pagesPathogenic Gram Positive BacteriaFahimRazmandehNo ratings yet
- Vacuna Hepatitis BDocument2 pagesVacuna Hepatitis Bluis alejandro rodriguez moraNo ratings yet
- The Biology of Skin ColorDocument4 pagesThe Biology of Skin ColorAntonov MikalisNo ratings yet
- Genetics Lecture 2 - DNA and Chromosome Structure PDFDocument58 pagesGenetics Lecture 2 - DNA and Chromosome Structure PDFMarlee SandersonNo ratings yet
- Activity ProposalSeminarDocument6 pagesActivity ProposalSeminarWil AlgonesNo ratings yet
- The Requirements of Mannose Binding LectinDocument2 pagesThe Requirements of Mannose Binding LectinNurul NadiaNo ratings yet
- FCE Practice Tests 1 2Document2 pagesFCE Practice Tests 1 2charlin.montalvanNo ratings yet