Download as pdf or txt
You might also like
- Hirac - DoleDocument11 pagesHirac - DoleAngela Salumbides50% (2)
- Sablin Et Al 2022 The Epigravettian Site of Yudinovo Russia Mammoth Bone Structures As Ritualised MiddensDocument22 pagesSablin Et Al 2022 The Epigravettian Site of Yudinovo Russia Mammoth Bone Structures As Ritualised MiddensEmerson NobreNo ratings yet
- Obsidian and Early Cultural Contact in The Near EastDocument43 pagesObsidian and Early Cultural Contact in The Near EastatheerNo ratings yet
- New AMS Dates From The Sub-Neolithic Sites in The Southern Buh Area (Ukraine) and Problems in The Buh-Dnister Culture ChronologyDocument30 pagesNew AMS Dates From The Sub-Neolithic Sites in The Southern Buh Area (Ukraine) and Problems in The Buh-Dnister Culture ChronologyDmytro_GaskevychNo ratings yet
- ПЖ Англ вариант PALJ445Document19 pagesПЖ Англ вариант PALJ445carlos.emr1991No ratings yet
- Functional Analysis of Late Holocene Flaked and PeDocument10 pagesFunctional Analysis of Late Holocene Flaked and PeCercea PaulaNo ratings yet
- LL - Studia Archeologica - 6 (SAUS)Document344 pagesLL - Studia Archeologica - 6 (SAUS)goshoNo ratings yet
- Recent Bivalve Ecology Applied To The Reconstruction of PaleocommunitiesDocument5 pagesRecent Bivalve Ecology Applied To The Reconstruction of PaleocommunitiesasmatiNo ratings yet
- Kocsis L. 2007 - Central Paratethyan Shark Fauna (Ipolytarnóc, Hungary)Document15 pagesKocsis L. 2007 - Central Paratethyan Shark Fauna (Ipolytarnóc, Hungary)Giovanni GalliNo ratings yet
- Poezii - M.eminescuDocument18 pagesPoezii - M.eminescuProciuc MarianaNo ratings yet
- Archeologia BalticaDocument16 pagesArcheologia BalticaMonika Gąsiorowska-DuszyńskaNo ratings yet
- Contextual Taphonomy of Worked Bones inDocument13 pagesContextual Taphonomy of Worked Bones inCarlos Andrés González ForeroNo ratings yet
- AJA, 91,1987 Chronology and Function of Ceramic UnguentariaDocument19 pagesAJA, 91,1987 Chronology and Function of Ceramic UnguentariaKarl HansNo ratings yet
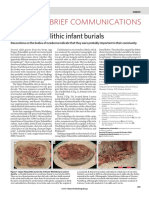
- Brief Communications: Upper Palaeolithic Infant BurialsDocument1 pageBrief Communications: Upper Palaeolithic Infant BurialsladuchessNo ratings yet
- Voichyshyn V. 2006b On The Revision of P.Balabai's Collection of Early Devonian Osteostraci (Agnatha) From Podolia (Ukraine) PDFDocument21 pagesVoichyshyn V. 2006b On The Revision of P.Balabai's Collection of Early Devonian Osteostraci (Agnatha) From Podolia (Ukraine) PDFvicNo ratings yet
- From Studies On Cultures of The 4TH Millennium B.C. in The Central Part of The Polish Coastal AreaDocument42 pagesFrom Studies On Cultures of The 4TH Millennium B.C. in The Central Part of The Polish Coastal AreaJustyna OrłowskaNo ratings yet
- Oravkinova Hromadova Vlaciky 2017 PDFDocument58 pagesOravkinova Hromadova Vlaciky 2017 PDFDominika OravkinováNo ratings yet
- Olorgesailie New Excavations and FindingDocument8 pagesOlorgesailie New Excavations and FindingGabriel MiguezNo ratings yet
- Bouzek Ondrejova 1988 Sindos Trebenishte Duvanli Interreations Between Thrace Macedonia and Greece in The 6th and 5th Centuries BC PDFDocument12 pagesBouzek Ondrejova 1988 Sindos Trebenishte Duvanli Interreations Between Thrace Macedonia and Greece in The 6th and 5th Centuries BC PDFОлеся КалашниковаNo ratings yet
- The Neolithic of The CaucasusDocument36 pagesThe Neolithic of The CaucasusNikoloz TsikaridzeNo ratings yet
- Artículo Acevedoetal2021Document11 pagesArtículo Acevedoetal2021LorenaNo ratings yet
- Pregled Puževa Golaća U Bokokotorskom Zaljevu, Crna Gora PDFDocument12 pagesPregled Puževa Golaća U Bokokotorskom Zaljevu, Crna Gora PDFKristina Noëlle TijanNo ratings yet
- Neolicization of AnatoliaDocument9 pagesNeolicization of AnatoliaTiago SochaczewskiNo ratings yet
- 49956323-Fraser Et Alii 2018Document11 pages49956323-Fraser Et Alii 2018Dai Tovar HellínNo ratings yet
- LV Kuce I Sahranjivanje EngleskiDocument20 pagesLV Kuce I Sahranjivanje EngleskiNenad TrajkovicNo ratings yet
- Bartosiewicz (2012) - Mammalian Remains From KorosDocument10 pagesBartosiewicz (2012) - Mammalian Remains From KorosDanaGrujićNo ratings yet
- Boric Price PNAS 2013 NBDocument7 pagesBoric Price PNAS 2013 NBIva TintiloNo ratings yet
- Individual Geographic Mobility in A Viking-Age Emporium-Burial Practices and Strontium Isotope Analyses of Ribe's Earliest InhabitantsDocument24 pagesIndividual Geographic Mobility in A Viking-Age Emporium-Burial Practices and Strontium Isotope Analyses of Ribe's Earliest InhabitantsBefore AfterNo ratings yet
- Mortuary Practices Bodies and Persons in The Neolithic and Early-Middle Copper Age of Southeast EuropeDocument34 pagesMortuary Practices Bodies and Persons in The Neolithic and Early-Middle Copper Age of Southeast EuropeRiad Ćišić100% (2)
- 2021 IAPO Bulletin 2021-2 2 SleptsovaDocument12 pages2021 IAPO Bulletin 2021-2 2 SleptsovaNećeš SadaNo ratings yet
- A Pilot Study of Black Chert Sourcing An PDFDocument9 pagesA Pilot Study of Black Chert Sourcing An PDF1heUndertakerNo ratings yet
- Obsidian Source Identification at Gre FillaDocument1 pageObsidian Source Identification at Gre FillaHalli PshdariNo ratings yet
- Jensen 2003 Trace Fossils A ReviewDocument10 pagesJensen 2003 Trace Fossils A ReviewGg GNo ratings yet
- Spondylus Gaederopus in Aegean PrehistorDocument14 pagesSpondylus Gaederopus in Aegean Prehistorpamant2No ratings yet
- Diversity of Desmid AlgaeDocument13 pagesDiversity of Desmid AlgaeLuisCarlosVillarrealDíazNo ratings yet
- Ancient Genomic Time Transect From The Central Asian Steppe Unravels The History of The ScythiansDocument15 pagesAncient Genomic Time Transect From The Central Asian Steppe Unravels The History of The Scythianscbk123No ratings yet
- Genome Sequence of A 45,000-Year-Old Modern Human From Western SiberiaDocument14 pagesGenome Sequence of A 45,000-Year-Old Modern Human From Western SiberiaPamela Rios Monteza100% (1)
- Ouranopithecus MacedoniensisDocument3 pagesOuranopithecus Macedoniensisstakis123No ratings yet
- Fossilrecordofangiosperms Nova 2012Document73 pagesFossilrecordofangiosperms Nova 2012Evaldo PapeNo ratings yet
- Journal of Archaeological Science: ReportsDocument8 pagesJournal of Archaeological Science: ReportsFilip OndrkálNo ratings yet
- Early Agricultural Colonisation of Peripheral Areas of Loess Uplands: New Data From Sandomierz Upland, PolandDocument8 pagesEarly Agricultural Colonisation of Peripheral Areas of Loess Uplands: New Data From Sandomierz Upland, Polandsnickercat.meowNo ratings yet
- Crvena Stijena Preliminary Report 2006-2009Document29 pagesCrvena Stijena Preliminary Report 2006-2009lord azraelNo ratings yet
- NewsletterMarch2007 SinglePage PDFDocument12 pagesNewsletterMarch2007 SinglePage PDFdtwigg123No ratings yet
- From The Sunghir Children To The Romito Dwarf - Aspects of The Upper Paleolithic Funerary LandscapeDocument8 pagesFrom The Sunghir Children To The Romito Dwarf - Aspects of The Upper Paleolithic Funerary LandscapeMaja100% (3)
- Alperson-Alfil 2008, Fire-Making at Gesher Benot Ya AqovDocument7 pagesAlperson-Alfil 2008, Fire-Making at Gesher Benot Ya AqovKevin López LealNo ratings yet
- New Ediacaran Fossils From The Ukraine, Some With A Putative Tunicate RelationshipDocument17 pagesNew Ediacaran Fossils From The Ukraine, Some With A Putative Tunicate RelationshipRoman MuzychenkoNo ratings yet
- Kovacs Gal & Bartosiewitz 2010 EN Hungary - IbranyDocument17 pagesKovacs Gal & Bartosiewitz 2010 EN Hungary - IbranyDanaGrujićNo ratings yet
- Ancient Genomic Time Transect From The Central Asian Steppe Unravels The History of The ScythiansDocument15 pagesAncient Genomic Time Transect From The Central Asian Steppe Unravels The History of The ScythiansErwin SchrödringerNo ratings yet
- Agiasmatsi: A Greek Cave Sanctuary in SphakiaDocument54 pagesAgiasmatsi: A Greek Cave Sanctuary in Sphakiaaguara01No ratings yet
- Atti Della Società Per La Preistoria e Protostoria Della Regione Friuli-Venezia Giulia, Volume XVII, 2008-2009. TriesteDocument200 pagesAtti Della Società Per La Preistoria e Protostoria Della Regione Friuli-Venezia Giulia, Volume XVII, 2008-2009. TriestePaolo BiagiNo ratings yet
- Some Statistics About Rock-Carvings of Lake Onega: Folklore (Estonia) October 1999Document11 pagesSome Statistics About Rock-Carvings of Lake Onega: Folklore (Estonia) October 1999Joel ArellanoNo ratings yet
- Biospeleological Bibliography of The Balkan PeninsulaDocument104 pagesBiospeleological Bibliography of The Balkan PeninsulaJozef DozsaiNo ratings yet
- Timargarha SkullsDocument147 pagesTimargarha SkullsHofmann SergioNo ratings yet
- Eastern Myth Elements in The West in LigDocument29 pagesEastern Myth Elements in The West in LigBogyeszArchNo ratings yet
- Ancient DNA Reveals Prehistoric Gene-Flow From Siberia in The Complex Human Population History of North East Europe - PLOS GeneticsDocument34 pagesAncient DNA Reveals Prehistoric Gene-Flow From Siberia in The Complex Human Population History of North East Europe - PLOS Geneticssariina.ghNo ratings yet
- Keyser 2009 PDFDocument16 pagesKeyser 2009 PDFkdNo ratings yet
- 1 s2.0 S1631068315000536 MainDocument11 pages1 s2.0 S1631068315000536 MainJustin SingletonNo ratings yet
- Australasian Fossils: A Students' Manual of PalaeontologyFrom EverandAustralasian Fossils: A Students' Manual of PalaeontologyNo ratings yet
- A New Genus of Pennsylvania Fish (Crossoperygii, Coelacanthiformes) from KansasFrom EverandA New Genus of Pennsylvania Fish (Crossoperygii, Coelacanthiformes) from KansasNo ratings yet
- Holocene Distribution and Extinction of The Moose (Alces Alces, Cervidae) in Central EuropeDocument17 pagesHolocene Distribution and Extinction of The Moose (Alces Alces, Cervidae) in Central EuropeDanaGrujićNo ratings yet
- FoliaArchaeologica 31 Pages55-75Document21 pagesFoliaArchaeologica 31 Pages55-75DanaGrujićNo ratings yet
- Climate Fluctuations and Trajectories ToDocument14 pagesClimate Fluctuations and Trajectories ToDanaGrujićNo ratings yet
- 186 1999 BerlinDiversitasDocument29 pages186 1999 BerlinDiversitasDanaGrujićNo ratings yet
- Krznarić-Škrivanko 2014 - SopotDocument37 pagesKrznarić-Škrivanko 2014 - SopotDanaGrujićNo ratings yet
- Animal PerceptionDocument11 pagesAnimal PerceptionDanaGrujićNo ratings yet
- Animal RightDocument15 pagesAnimal RightDanaGrujićNo ratings yet
- Animal ExtinctionsDocument20 pagesAnimal ExtinctionsDanaGrujićNo ratings yet
- Bogaard (2005) - Garden AgricultureDocument21 pagesBogaard (2005) - Garden AgricultureDanaGrujićNo ratings yet
- Review Articles: Spiritual Habitus and The Archaeology of BeliefDocument24 pagesReview Articles: Spiritual Habitus and The Archaeology of BeliefDanaGrujićNo ratings yet
- Kreuz Et Al. (2020) - Hungarian Neolithic Landscapes CropsDocument17 pagesKreuz Et Al. (2020) - Hungarian Neolithic Landscapes CropsDanaGrujićNo ratings yet
- Sacred Landscapes of Siberia Symbolic Uses of SpacDocument5 pagesSacred Landscapes of Siberia Symbolic Uses of SpacDanaGrujićNo ratings yet
- Living With Water in Vojvodina: Andjelka Belic, Radovan Sa Vic & Si Ma BelicDocument6 pagesLiving With Water in Vojvodina: Andjelka Belic, Radovan Sa Vic & Si Ma BelicDanaGrujićNo ratings yet
- Vertebrate Fauna at The Neolithic and Eneolithic S PDFDocument12 pagesVertebrate Fauna at The Neolithic and Eneolithic S PDFDanaGrujićNo ratings yet
- Barker (1975) - Early Neo Landuse in YUDocument20 pagesBarker (1975) - Early Neo Landuse in YUDanaGrujićNo ratings yet
- Crane Food Pet and SymbolDocument12 pagesCrane Food Pet and SymbolDanaGrujićNo ratings yet
- The Ratio of Domestic and Wild Animals at NeolithiDocument9 pagesThe Ratio of Domestic and Wild Animals at NeolithiDanaGrujićNo ratings yet
- Animal Husbandry and Hunting in Roman Provinces in SerbiaDocument13 pagesAnimal Husbandry and Hunting in Roman Provinces in SerbiaDanaGrujićNo ratings yet
- Erdogu 2014 GOKCEADA - UGURLU Field Season 2011-2013Document22 pagesErdogu 2014 GOKCEADA - UGURLU Field Season 2011-2013DanaGrujićNo ratings yet
- A Leopard Panthera Pardus L 1758 Find FRDocument11 pagesA Leopard Panthera Pardus L 1758 Find FRDanaGrujićNo ratings yet
- Radmanović Et Al. (2015) - Wild-Domestic VojvodinaDocument8 pagesRadmanović Et Al. (2015) - Wild-Domestic VojvodinaDanaGrujićNo ratings yet
- Boric. Gurova, Miracle PDFDocument7 pagesBoric. Gurova, Miracle PDFDanaGrujićNo ratings yet
- Radmanović Et Al. Ornitofauna VojvodinaDocument8 pagesRadmanović Et Al. Ornitofauna VojvodinaDanaGrujićNo ratings yet
- Environmental Stress in Early Domestic SDocument11 pagesEnvironmental Stress in Early Domestic SDanaGrujićNo ratings yet
- Bartosiewicz (2012) - Mammalian Remains From KorosDocument10 pagesBartosiewicz (2012) - Mammalian Remains From KorosDanaGrujićNo ratings yet
- Bartosiewicz (2012) - Fish Remains From Koros PDFDocument6 pagesBartosiewicz (2012) - Fish Remains From Koros PDFDanaGrujićNo ratings yet
- Minimum Cost Spanning TreesDocument4 pagesMinimum Cost Spanning TreesSahal CSNo ratings yet
- A Comparative Study Chap1 5Document40 pagesA Comparative Study Chap1 5Mark Joshua MariñasNo ratings yet
- Definitive Budget of Work For Sy 2021-2022 (Languages) : English 6Document4 pagesDefinitive Budget of Work For Sy 2021-2022 (Languages) : English 6catherine renanteNo ratings yet
- Module 2 - Form, Skin and ClimateDocument15 pagesModule 2 - Form, Skin and ClimateĐặng Gia HoàngNo ratings yet
- Solid Liquid Filtration and Separation Technology - 1996 - Rushton - Appendix A Particle Size Shape and SizeDocument15 pagesSolid Liquid Filtration and Separation Technology - 1996 - Rushton - Appendix A Particle Size Shape and SizeDevpriy sahuNo ratings yet
- What Are Spring and Neap TidesDocument10 pagesWhat Are Spring and Neap TidesChris LibariosNo ratings yet
- Solutions To Selected Exercises: Categorical Data Analysis, 3Rd EditionDocument30 pagesSolutions To Selected Exercises: Categorical Data Analysis, 3Rd EditionLiseNo ratings yet
- Given Three Points Compute Affine Transformation - Stack OverflowDocument1 pageGiven Three Points Compute Affine Transformation - Stack OverflowWichoNo ratings yet
- Catalogue 30XW-V Water Cooled Screw InverterDocument26 pagesCatalogue 30XW-V Water Cooled Screw Inverterekoindri.rkh2022No ratings yet
- Activity 7 Assessment 7Document9 pagesActivity 7 Assessment 7Shey FuentesNo ratings yet
- Plan de InglésDocument25 pagesPlan de InglésAlejandro Zuleta GarciaNo ratings yet
- GK 1 1Document7 pagesGK 1 1Muhammad AliNo ratings yet
- And In-Depth Analysis To Score Full Credit.: Spring 2020 ISOM3730 Final Exam SolutionDocument5 pagesAnd In-Depth Analysis To Score Full Credit.: Spring 2020 ISOM3730 Final Exam SolutionSin TungNo ratings yet
- D.I.S.S Report (Bonifacio)Document16 pagesD.I.S.S Report (Bonifacio)Lovely CarbonelNo ratings yet
- 190102-DC10.5-Expansive and Stable Soils-Public ReviewDocument66 pages190102-DC10.5-Expansive and Stable Soils-Public Reviewvõ hoàng thái50% (2)
- 4 Continuous Probability Distribution.9188.1578362393.1974Document47 pages4 Continuous Probability Distribution.9188.1578362393.1974Pun AditepNo ratings yet
- P.1 Biasedness - The Bias of On Estimator Is Defined As:: Chapter Two EstimatorsDocument8 pagesP.1 Biasedness - The Bias of On Estimator Is Defined As:: Chapter Two EstimatorsFerekkanNo ratings yet
- MSDS NaOH 0.1 NDocument5 pagesMSDS NaOH 0.1 NYoga Maulana RahmanNo ratings yet
- Full Download PDF of (Ebook PDF) Calculus & Its Applications, Brief Version 14th Edition All ChapterDocument40 pagesFull Download PDF of (Ebook PDF) Calculus & Its Applications, Brief Version 14th Edition All Chaptersimthebutana100% (11)
- Geological Hazard and Engineering Geological Map of DilaDocument140 pagesGeological Hazard and Engineering Geological Map of DilaMowlid MohammudNo ratings yet
- Astm D5391 - 23Document8 pagesAstm D5391 - 23Bay NguyenNo ratings yet
- Tensile TestDocument22 pagesTensile TestFlying KiteNo ratings yet
- Basic Listening and Speaking With AudioDocument36 pagesBasic Listening and Speaking With AudiokamalNo ratings yet
- National Wind Farm Development GuidelinesDocument208 pagesNational Wind Farm Development GuidelinesYara Abu DakkaNo ratings yet
- Lesson Plan Class 3 English Chapter 6 To 12Document60 pagesLesson Plan Class 3 English Chapter 6 To 12Tamuna AkhobadzeNo ratings yet
- A Faithful Solid-State Spin-Wave Quantum Memory For Polarization QubitsDocument8 pagesA Faithful Solid-State Spin-Wave Quantum Memory For Polarization QubitsJay-R Notorio PallegaNo ratings yet
- 1beginners English Book PDFDocument140 pages1beginners English Book PDFMarcelo Valdivieso rojas100% (1)
- SCS SF Self Compassion Scale Blank Form PDFDocument1 pageSCS SF Self Compassion Scale Blank Form PDFAurooba N.No ratings yet
- STS 5 Indigenous Science and Technology in The PhilDocument12 pagesSTS 5 Indigenous Science and Technology in The PhilTheo SeriyeNo ratings yet