Download as pdf or txt
You might also like
- Iso 15213-1-2023Document28 pagesIso 15213-1-2023JOSEPH100% (7)
- Legend of The Arch Magus - Curse of Agares Volume 5Document178 pagesLegend of The Arch Magus - Curse of Agares Volume 5TgnNo ratings yet
- Checklist Audit ISO 22000Document18 pagesChecklist Audit ISO 22000Abhishek Kumar Singh95% (20)
- Kiss NotesDocument38 pagesKiss NotesNalini Prakash100% (2)
- Trim Clean 2030-Msds (Bld-1656 v1)Document8 pagesTrim Clean 2030-Msds (Bld-1656 v1)Tri Febrianto PamungkasNo ratings yet
- A New Specimen of Sinosaurus Triassicus Dinosauria Theropoda From The Early Jurassic of Lufeng Yunnan ChinaDocument16 pagesA New Specimen of Sinosaurus Triassicus Dinosauria Theropoda From The Early Jurassic of Lufeng Yunnan Chinamauricio.garciaNo ratings yet
- Hatchlings of The Deep-Sea Octopus Graneledone Boreopacifica Are The Largest and Most Advanced KnownDocument3 pagesHatchlings of The Deep-Sea Octopus Graneledone Boreopacifica Are The Largest and Most Advanced KnownCarlos MeirellesNo ratings yet
- Viviparity and K-Selected Life History in A Mesozoic Marine Plesiosaur Reptilia SauropterygiaDocument15 pagesViviparity and K-Selected Life History in A Mesozoic Marine Plesiosaur Reptilia SauropterygianoaraptorNo ratings yet
- Ancient DNA Reveals Monozygotic Newborn TwinsDocument11 pagesAncient DNA Reveals Monozygotic Newborn TwinsОльга ПіхуляNo ratings yet
- TIBURONº1Document21 pagesTIBURONº1Jhasmani AmcNo ratings yet
- 2011 Tsuihiji Et Al. Tarbosaurus Juvenile Skull PROOFDocument21 pages2011 Tsuihiji Et Al. Tarbosaurus Juvenile Skull PROOFKanekoNo ratings yet
- J Ijpp 2012 10 002Document8 pagesJ Ijpp 2012 10 002Pinkky PiNo ratings yet
- Comparitive Anatomy BipedalismDocument16 pagesComparitive Anatomy BipedalismGeorge RyleNo ratings yet
- Enigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDocument6 pagesEnigmatic Amphibians in Mid-Cretaceous Amber Were Chameleon-Like Ballistic FeedersDirtyBut WholeNo ratings yet
- Sexual Dimorphism and Life History of Keichousaurus Hui (Reptilia: Sauropterygia)Document9 pagesSexual Dimorphism and Life History of Keichousaurus Hui (Reptilia: Sauropterygia)Adnana CebucNo ratings yet
- Evolutionary History of Falkland Wolf (Slater Et Al)Document2 pagesEvolutionary History of Falkland Wolf (Slater Et Al)Francisco J. OvalleNo ratings yet
- Right Handed Neandertals: Vindija and BeyondDocument16 pagesRight Handed Neandertals: Vindija and BeyondTabulastorNo ratings yet
- Joa 13363Document15 pagesJoa 13363EdisonNo ratings yet
- Genome Sequence of A 45,000-Year-Old Modern Human From Western SiberiaDocument14 pagesGenome Sequence of A 45,000-Year-Old Modern Human From Western SiberiaPamela Rios Monteza100% (1)
- GauthierDocument306 pagesGauthierDan IstrateNo ratings yet
- Ward 2015 AustralopitecinosDocument14 pagesWard 2015 Australopitecinostomascostelafre1410No ratings yet
- Hublin Et Al 2017 African Origin H Sapiens MoroccoDocument17 pagesHublin Et Al 2017 African Origin H Sapiens MoroccoRodrigo Giesta FigueiredoNo ratings yet
- Nature 22336Document18 pagesNature 22336Elmer L'éléphantNo ratings yet
- SullivanetalDocument13 pagesSullivanetalDianitaRocNo ratings yet
- Chhetup, Human Skeleton, Australia, PaleolithicDocument8 pagesChhetup, Human Skeleton, Australia, PaleolithicVolinthosNo ratings yet
- Casey Anthony - UF Skeletal Dispersal ReportDocument10 pagesCasey Anthony - UF Skeletal Dispersal ReportTruthTronNo ratings yet
- Brochu - EXTINCT MALAGASY CROCODILEDocument29 pagesBrochu - EXTINCT MALAGASY CROCODILECrocodilo VasconcelosNo ratings yet
- Wbiea 1765Document17 pagesWbiea 1765ooksa GamingNo ratings yet
- First Pleistocene Jumping Mouse Zapus ZapodinaeDocument7 pagesFirst Pleistocene Jumping Mouse Zapus Zapodinaelophostoma1No ratings yet
- Lab9 AustralopithecineReading F17Document12 pagesLab9 AustralopithecineReading F17WeaselmonkeyNo ratings yet
- Radiometric Dating of The Type-Site For Homo Heidelbergensis at Mauer, GermanyDocument5 pagesRadiometric Dating of The Type-Site For Homo Heidelbergensis at Mauer, GermanylapocheqseachipartioNo ratings yet
- The Lower Jurassic Ornithischian Dinosaur Cranial Anatomy, Functional Morphology, Taxonomy, and RelationshipsDocument95 pagesThe Lower Jurassic Ornithischian Dinosaur Cranial Anatomy, Functional Morphology, Taxonomy, and Relationshipsbraydenstoch777No ratings yet
- The Origin of The Turtle Body Plan EvideDocument19 pagesThe Origin of The Turtle Body Plan EvideXweuis Hekuos KweNo ratings yet
- 10 1016@j Geobios 2020 09 005Document36 pages10 1016@j Geobios 2020 09 005DirtyBut WholeNo ratings yet
- Bambiraptor and DeinonychusDocument10 pagesBambiraptor and DeinonychusFelipe MesVasNo ratings yet
- Perry Et Al. 2014Document15 pagesPerry Et Al. 2014Maria Elena Chavez MarreroNo ratings yet
- Five Years of H Flores Iens IsDocument13 pagesFive Years of H Flores Iens IsnomadNo ratings yet
- Climbing AstroblepusDocument16 pagesClimbing AstroblepusluisaNo ratings yet
- 10.1007@s10641 017 0668 0Document6 pages10.1007@s10641 017 0668 0Marina Trillo GabaldonNo ratings yet
- A Synopsis of Neotropical Hylid Frogs, Genus OsteocephalusFrom EverandA Synopsis of Neotropical Hylid Frogs, Genus OsteocephalusNo ratings yet
- App44 189Document31 pagesApp44 189RoullienNo ratings yet
- Soft Anatomy of The Early CambrianDocument10 pagesSoft Anatomy of The Early CambrianEdna AlvarezNo ratings yet
- Yan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaDocument12 pagesYan Et Al. 2014. The Most Mysterious Beetles Jurassic Jurodidae (Insecta Coleoptera) From ChinaCarlos A. JulioNo ratings yet
- Some Observations On The Habits and Placentation of Tatu NovemcinctumDocument110 pagesSome Observations On The Habits and Placentation of Tatu NovemcinctumcerradointernetNo ratings yet
- Ciclostomi FosiliDocument5 pagesCiclostomi FosiliAnca MihalcescuNo ratings yet
- Highlights of Cetacean Embryology: J. G. M. ThewissenDocument12 pagesHighlights of Cetacean Embryology: J. G. M. ThewissenFrancisco Hernandez DuqueNo ratings yet
- Nesbitt Et Al 2023 Mambachiton Fiandohana The Earliest-Diverging AvemetatarsalianDocument28 pagesNesbitt Et Al 2023 Mambachiton Fiandohana The Earliest-Diverging Avemetatarsalianmauricio.garciaNo ratings yet
- Kenyanthropus PlatyopsDocument4 pagesKenyanthropus Platyopsjohnny cartinNo ratings yet
- Experimental Study of Bone Modification by Captive CaracalDocument13 pagesExperimental Study of Bone Modification by Captive CaracalSidra ZahoorNo ratings yet
- Australopithecus Phylogeny H. SellassieDocument9 pagesAustralopithecus Phylogeny H. SellassiepgandzNo ratings yet
- Kostick, 1963Document12 pagesKostick, 1963t0limanNo ratings yet
- Patterns of Activity-Induced Pathology in a Canadian Inuit PopulationFrom EverandPatterns of Activity-Induced Pathology in a Canadian Inuit PopulationNo ratings yet
- The Appendicular Skeleton of Suuwassea eDocument22 pagesThe Appendicular Skeleton of Suuwassea enomadNo ratings yet
- Morphology of The Caudal Vertebrae in RHDocument15 pagesMorphology of The Caudal Vertebrae in RHMIGUEL ANGEL AVILA GUERRANo ratings yet
- IESO Preparation Material (Paleontology)Document25 pagesIESO Preparation Material (Paleontology)Science Olympiad Blog67% (3)
- Mother Tongue 14 (August 1991)Document65 pagesMother Tongue 14 (August 1991)Allan BomhardNo ratings yet
- Lyson Etal 2013 Evolutionary Origin of The Turtle ShellDocument7 pagesLyson Etal 2013 Evolutionary Origin of The Turtle ShellThiago S. SalesNo ratings yet
- Biostratigraphy of The Scalenodontoides Assemblage Zone (Stormberg Group, Karoo Supergroup), South AfricaDocument10 pagesBiostratigraphy of The Scalenodontoides Assemblage Zone (Stormberg Group, Karoo Supergroup), South AfricaNeo MetsingNo ratings yet
- Craniomandibular Anatomy of A Juvenile Specimen of Edmontosaurus Regalis Lambe 1917 Clarifies Issues in Ontogeny and BiogeographyDocument22 pagesCraniomandibular Anatomy of A Juvenile Specimen of Edmontosaurus Regalis Lambe 1917 Clarifies Issues in Ontogeny and BiogeographyCrocodilo VasconcelosNo ratings yet
- Farm Beneath The SandDocument15 pagesFarm Beneath The SandwaxwonderNo ratings yet
- Revision of The Unique Early Cretaceous Mecoptera From Koonwarra (Australia) With Description of A New Genus and FamilyDocument6 pagesRevision of The Unique Early Cretaceous Mecoptera From Koonwarra (Australia) With Description of A New Genus and Familyagnieszka.soszynska.majNo ratings yet
- 2019 - Chen - Trilobate Bilaterian Elucidates Early Animal EvolutionDocument16 pages2019 - Chen - Trilobate Bilaterian Elucidates Early Animal EvolutionDredeltoNo ratings yet
- 3441 - 407803 - Sangiran Bailey2014Document9 pages3441 - 407803 - Sangiran Bailey2014TyasNo ratings yet
- GCSE-Biology-7-Summer2019-Foundation Tier, Unit 1-PaperDocument24 pagesGCSE-Biology-7-Summer2019-Foundation Tier, Unit 1-PaperjramatlhakolaneNo ratings yet
- Filter BrochureDocument2 pagesFilter BrochureCanadaPool100% (1)
- Observations From The Plant VisitDocument2 pagesObservations From The Plant VisitAniket ShrivastavaNo ratings yet
- Kisi - Kisi LKS Jateng 2018 - AssemblyDocument27 pagesKisi - Kisi LKS Jateng 2018 - AssemblyZidNa Munif MuhammadNo ratings yet
- Repro enDocument20 pagesRepro enWai Mei TeeNo ratings yet
- Performance KPI's Optimization TipsDocument17 pagesPerformance KPI's Optimization TipsShekh Mohammad NasimNo ratings yet
- Uasin Gishu - 2Document77 pagesUasin Gishu - 2peeteoNo ratings yet
- 5th Fancy BreadDocument7 pages5th Fancy BreadMera Funportal0% (1)
- Makita 6910Document3 pagesMakita 6910El MacheteNo ratings yet
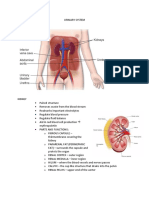
- Urinary SystemDocument9 pagesUrinary SystemMary Joyce RamosNo ratings yet
- Food Menu TemplateDocument14 pagesFood Menu TemplateG- WizNo ratings yet
- Philips PL - 2016Document52 pagesPhilips PL - 2016Ganesh PonmozhiNo ratings yet
- BS Soil and Rock Explanatory NotesDocument15 pagesBS Soil and Rock Explanatory NotesvictorNo ratings yet
- Maya Under Water LightingDocument12 pagesMaya Under Water LightingKombiah RkNo ratings yet
- 19 - Bearing Capacity - Eccentric LoadingDocument5 pages19 - Bearing Capacity - Eccentric LoadinghiyeonNo ratings yet
- Wollo University: Kombolcha Institute of TechnologyDocument12 pagesWollo University: Kombolcha Institute of Technologymike workuNo ratings yet
- Stalking & Dreaming PDFDocument4 pagesStalking & Dreaming PDFJoannaAllen100% (2)
- Plag - ReportDocument6 pagesPlag - ReportMehedi HasanNo ratings yet
- Pathway - Skoliosis GROUPDocument12 pagesPathway - Skoliosis GROUPAnonymous NZTQVgjaNo ratings yet
- Assignment2 NamocDocument5 pagesAssignment2 NamocHenry Darius NamocNo ratings yet
- Experiment 5Document21 pagesExperiment 5Jervin JamillaNo ratings yet
- Paper 2222Document16 pagesPaper 2222Abhijeet GholapNo ratings yet
- Covellite PDFDocument1 pageCovellite PDFRyoga RizkyNo ratings yet
- Novilyn C. Pataray BSN - Ii Thalassemia Major: St. Paul College of Ilocos SurDocument1 pageNovilyn C. Pataray BSN - Ii Thalassemia Major: St. Paul College of Ilocos SurCharina Aubrey0% (1)
- Maersk IntrepidDocument8 pagesMaersk Intrepidrickyngs100% (1)
- Environmental EthicsDocument6 pagesEnvironmental EthicsHELI SHAHNo ratings yet