
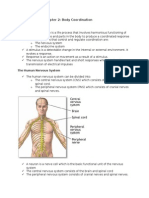
Chapter II Module
Chapter II Module
You might also like
- Unit 3 Module 1Document6 pagesUnit 3 Module 1heide100% (1)
- Zen Shiatsu III Kyo/jitsu Study Guide: General Metal Healthy FunctionsDocument13 pagesZen Shiatsu III Kyo/jitsu Study Guide: General Metal Healthy FunctionsbigkeitasNo ratings yet
- PallorDocument16 pagesPallorManal AlQuaimi100% (1)
- S6 BIO (Nervous Coordination) Part2Document35 pagesS6 BIO (Nervous Coordination) Part2ivanpatrick658No ratings yet
- Blue and Cream Vintage General ProposalDocument12 pagesBlue and Cream Vintage General ProposalKimberly Jane MitraNo ratings yet
- CAPITIN - AnSci1.1 Anatomy and Physiology of Farm AnimalsDocument6 pagesCAPITIN - AnSci1.1 Anatomy and Physiology of Farm AnimalsJuan CapitinNo ratings yet
- Chapter 3Document7 pagesChapter 3Azzem JenalNo ratings yet
- CompilationDocument63 pagesCompilationJenny V. BajoNo ratings yet
- Nervous System and Coordination of Nervous and Endocrine System To Achieve HomeostasisDocument65 pagesNervous System and Coordination of Nervous and Endocrine System To Achieve HomeostasisJoseph yvannNo ratings yet
- How Hormones Govern Body ActivitiesDocument4 pagesHow Hormones Govern Body ActivitiesMarvin MelisNo ratings yet
- Esson 3 Feedback Mechanism and Homeostasis StudentDocument27 pagesEsson 3 Feedback Mechanism and Homeostasis StudentJean Marc ReyNo ratings yet
- Topic 13 John Paul Magtalas G BioDocument39 pagesTopic 13 John Paul Magtalas G BioRaon, Simone AndreiNo ratings yet
- NegOr Q4 GenBio2 SLKWeek4 v2Document20 pagesNegOr Q4 GenBio2 SLKWeek4 v2Maribel Ruedas RamosNo ratings yet
- Control and Coordination CL XDocument7 pagesControl and Coordination CL XMukul BanoulaNo ratings yet
- AnscieDocument10 pagesAnscieEstephanie BusligNo ratings yet
- MODULE 2 Ansci 1Document31 pagesMODULE 2 Ansci 1Nelgine GepuitNo ratings yet
- Science 10 Handout - Nervous and EndocrineDocument2 pagesScience 10 Handout - Nervous and EndocrineAngelica MartinNo ratings yet
- Activity 1 Animal SciDocument5 pagesActivity 1 Animal SciKylle BedisNo ratings yet
- Anatomy Fuj JobbyDocument15 pagesAnatomy Fuj JobbyObadah AbdulsattarNo ratings yet
- Endocrine System 1 PDFDocument10 pagesEndocrine System 1 PDFDilNo ratings yet
- Control and Coordination - Class XDocument5 pagesControl and Coordination - Class XMonika Mehan89% (19)
- Topic 3 - Nervous Control in Plants and AnimalsDocument5 pagesTopic 3 - Nervous Control in Plants and AnimalsYara Jeah Ingay BaliliNo ratings yet
- ABPG1103 - Topic 2 - Biological Psychology - 222Document31 pagesABPG1103 - Topic 2 - Biological Psychology - 222nureryani bathowiNo ratings yet
- Q3 - Nervous and Endocrine Systems - Science10 PDFDocument84 pagesQ3 - Nervous and Endocrine Systems - Science10 PDFGabriel Pasumbal100% (1)
- TVBS Nervous SystemDocument20 pagesTVBS Nervous SystemAngel Mae Masa FloresNo ratings yet
- Animal Organ Systems and Homeostasis PDFDocument13 pagesAnimal Organ Systems and Homeostasis PDFAnonymous HXLczq3No ratings yet
- Chapter 3Document109 pagesChapter 3amala raviNo ratings yet
- The Receiving Mechanism: Sense Organs Are Sensitive Nerve Endings Located in Certain Body Parts. They Are Receptors ofDocument5 pagesThe Receiving Mechanism: Sense Organs Are Sensitive Nerve Endings Located in Certain Body Parts. They Are Receptors ofArlene Cerdeña SalcedaNo ratings yet
- Science 10 Quarter 3 Module 3Document15 pagesScience 10 Quarter 3 Module 3Zahra Alexä DavidsonNo ratings yet
- B Science 10 Quarter 3 Module 2 (Week 3)Document17 pagesB Science 10 Quarter 3 Module 2 (Week 3)Daisy Soriano PrestozaNo ratings yet
- NCERT Summary Control & CoordinationDocument12 pagesNCERT Summary Control & Coordinationpadhako.harshNo ratings yet
- Chapter 1 - The Science of Anatomy and Physiology UDocument5 pagesChapter 1 - The Science of Anatomy and Physiology UJess SommervilleNo ratings yet
- Essay B. InggrisDocument9 pagesEssay B. InggrisM.HalilNo ratings yet
- Chapter 7 Bio Class 10Document15 pagesChapter 7 Bio Class 10Aysha KhanNo ratings yet
- Chemical and Nervous SystemDocument42 pagesChemical and Nervous SystemMa Divina Kristi DiscarNo ratings yet
- Psychology PresentationDocument16 pagesPsychology PresentationFasih RehmanNo ratings yet
- Control and Coordination Do You Know Which Organs Make Up The Nervous System?Document22 pagesControl and Coordination Do You Know Which Organs Make Up The Nervous System?So TemptedNo ratings yet
- Class 10 Chapter 7 NotesDocument7 pagesClass 10 Chapter 7 NotesIra LaratteNo ratings yet
- Nervous SystemDocument4 pagesNervous SystemTherese MaleriadoNo ratings yet
- What Is Human Endocrine SystemDocument38 pagesWhat Is Human Endocrine SystemNatukunda DianahNo ratings yet
- Control Coordination NotesDocument6 pagesControl Coordination NotesRaman ChaudharyNo ratings yet
- Discuss and Elaborate The Relationship of The Different Systems Found in The Animal BodyDocument3 pagesDiscuss and Elaborate The Relationship of The Different Systems Found in The Animal BodyJenny V. BajoNo ratings yet
- Cbse Class 10 Science Notes Chapter 7 Control and CoordinationDocument8 pagesCbse Class 10 Science Notes Chapter 7 Control and CoordinationFatema EshrakNo ratings yet
- Control and CoordinationDocument8 pagesControl and CoordinationkkNo ratings yet
- Unit 4 Perception and Coordination 1Document84 pagesUnit 4 Perception and Coordination 1Iker anubisNo ratings yet
- Control and CoordinationDocument11 pagesControl and CoordinationJit AggNo ratings yet
- Control and Coordination: - Prepared by-R.S.Chandanshiv, Principal, JNV, N.C.Hills (Assam)Document30 pagesControl and Coordination: - Prepared by-R.S.Chandanshiv, Principal, JNV, N.C.Hills (Assam)Sheetal Kendre DagadkhairNo ratings yet
- Fall 2022 - ZOO403 - 1 - BC190408115Document4 pagesFall 2022 - ZOO403 - 1 - BC190408115Misbah NoreenNo ratings yet
- Copy-REGULATORY SYSTEM HODocument8 pagesCopy-REGULATORY SYSTEM HORio FirmansyahNo ratings yet
- Week 19 HSBDocument15 pagesWeek 19 HSBedhanassarNo ratings yet
- Form 4 Science Chapter 2Document13 pagesForm 4 Science Chapter 2Cheah Foo Kit50% (2)
- Anatomy and Physiology ReviewerDocument10 pagesAnatomy and Physiology ReviewerMae Christelle FigueroaNo ratings yet
- Preparation in Psychology: Bicol University College of Engineering East Campus LegazpiDocument8 pagesPreparation in Psychology: Bicol University College of Engineering East Campus LegazpiKielNo ratings yet
- Anatomy of The BrainDocument38 pagesAnatomy of The BrainnayanaNo ratings yet
- Departemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraDocument21 pagesDepartemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraFira TasyaNo ratings yet
- Anatomy and Fisiology NoteDocument3 pagesAnatomy and Fisiology NoteIssac LauNo ratings yet
- NScELEC3 WEEK 9 LESSONDocument9 pagesNScELEC3 WEEK 9 LESSONAlyssa Jane AbellonNo ratings yet
- A. MedStu - CNSDocument20 pagesA. MedStu - CNSNwraz AlsahibNo ratings yet
- Nervous System FunctionsDocument10 pagesNervous System FunctionsJovi Leo PunoNo ratings yet
- Departemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraDocument20 pagesDepartemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraabbyNo ratings yet
- Gale Researcher Guide for: Overview of Physiology and NeuropsychologyFrom EverandGale Researcher Guide for: Overview of Physiology and NeuropsychologyNo ratings yet
- Intro To Human Anatomy & Physiology: Quick Review Notes Chapter 1From EverandIntro To Human Anatomy & Physiology: Quick Review Notes Chapter 1No ratings yet
- Chapter 2 UNIT IVDocument24 pagesChapter 2 UNIT IVAyen LatosaNo ratings yet
- Chapter 2 UNIT IIDocument10 pagesChapter 2 UNIT IIAyen LatosaNo ratings yet
- Chapter 2 UNIT 1Document19 pagesChapter 2 UNIT 1Ayen LatosaNo ratings yet
- Cardiovascular System AnatomyDocument54 pagesCardiovascular System AnatomyAyen LatosaNo ratings yet
- Blood Pressure - Blood Pressure ChartDocument3 pagesBlood Pressure - Blood Pressure ChartMahmoud MaherNo ratings yet
- Activity 8 - Blood and HematopoiesisDocument8 pagesActivity 8 - Blood and HematopoiesisMarissa CordovaNo ratings yet
- Ex 5 Activity 3Document4 pagesEx 5 Activity 3Rafeik05100% (2)
- Name - Soni Ritika Rajesh, Group - GM20-054-2A Physiology Week 03 TasksDocument10 pagesName - Soni Ritika Rajesh, Group - GM20-054-2A Physiology Week 03 TasksRitika SoniNo ratings yet
- Hemophilia: Joanna Marie de Guzman RNDocument35 pagesHemophilia: Joanna Marie de Guzman RNJoan VenidaNo ratings yet
- Ineffective Tissue PerfussionDocument2 pagesIneffective Tissue Perfussionmonico39No ratings yet
- Blood Groups SystemsDocument7 pagesBlood Groups SystemsMir jaliesNo ratings yet
- 【Zybio】Z3 Service Training PPT-Distributor 20200110Document116 pages【Zybio】Z3 Service Training PPT-Distributor 20200110Fermin Tafur Lliuya100% (1)
- Disciplina AquasDocument84 pagesDisciplina AquasMoises TellezNo ratings yet
- 7A Cells, Tissues and Organs Booklet - Doc-1Document16 pages7A Cells, Tissues and Organs Booklet - Doc-1Majdoline SadeddineNo ratings yet
- Neonatal Resuscitation PDFDocument1 pageNeonatal Resuscitation PDFOdessa EnriquezNo ratings yet
- Management and Prognosis of The Thalassemias - UpToDateDocument45 pagesManagement and Prognosis of The Thalassemias - UpToDatesushi37No ratings yet
- 1 Hema LecDocument5 pages1 Hema LecAngela ReyesNo ratings yet
- Basic Science JSS2 Question BankDocument3 pagesBasic Science JSS2 Question BankIsaac100% (1)
- Cardiogenic Shock NCLEX Review QuizDocument6 pagesCardiogenic Shock NCLEX Review QuizRegie Marie EvangelistaNo ratings yet
- Exploring Science Edition © Pearson Education Limited 2008Document2 pagesExploring Science Edition © Pearson Education Limited 2008Victor Barber Sanchis100% (1)
- Blood Banking Course BookDocument2 pagesBlood Banking Course BookShukr Wesman BlbasNo ratings yet
- Natural Eye CareDocument222 pagesNatural Eye Careskribdot100% (3)
- The Effects of Lifestyle To Respiratory and Circulatory SystemsDocument11 pagesThe Effects of Lifestyle To Respiratory and Circulatory SystemsSharalyn Sia100% (1)
- Circulatory and Respiratory System, Working TogetherDocument32 pagesCirculatory and Respiratory System, Working TogetherRhona AngelaNo ratings yet
- S4 Mock 1 2006 Biology P1Document8 pagesS4 Mock 1 2006 Biology P1cyber sec100% (1)
- Ha&P - Semi ReviewerDocument71 pagesHa&P - Semi ReviewerLittle StuartNo ratings yet
- Vox Sanguin Maret 2022 - OptDocument192 pagesVox Sanguin Maret 2022 - Optrsdarsono labNo ratings yet
- Kaigo Prometric Lesson (Eng)Document108 pagesKaigo Prometric Lesson (Eng)Ysmech SalazarNo ratings yet
- Iron MetabolismDocument3 pagesIron MetabolismlucaNo ratings yet
- Isaac Typhoid NegativeDocument5 pagesIsaac Typhoid Negativeganesh khondeNo ratings yet
- Blood Gas HandbookDocument112 pagesBlood Gas HandbookNuno Silva100% (1)
- Hkcee Biology - 4.6 Transport in Humans - P.1Document13 pagesHkcee Biology - 4.6 Transport in Humans - P.1irisyyy27No ratings yet





























































































Download as docx, pdf, or txt
You might also like
- Unit 3 Module 1Document6 pagesUnit 3 Module 1heide100% (1)
- Zen Shiatsu III Kyo/jitsu Study Guide: General Metal Healthy FunctionsDocument13 pagesZen Shiatsu III Kyo/jitsu Study Guide: General Metal Healthy FunctionsbigkeitasNo ratings yet
- PallorDocument16 pagesPallorManal AlQuaimi100% (1)
- S6 BIO (Nervous Coordination) Part2Document35 pagesS6 BIO (Nervous Coordination) Part2ivanpatrick658No ratings yet
- Blue and Cream Vintage General ProposalDocument12 pagesBlue and Cream Vintage General ProposalKimberly Jane MitraNo ratings yet
- CAPITIN - AnSci1.1 Anatomy and Physiology of Farm AnimalsDocument6 pagesCAPITIN - AnSci1.1 Anatomy and Physiology of Farm AnimalsJuan CapitinNo ratings yet
- Chapter 3Document7 pagesChapter 3Azzem JenalNo ratings yet
- CompilationDocument63 pagesCompilationJenny V. BajoNo ratings yet
- Nervous System and Coordination of Nervous and Endocrine System To Achieve HomeostasisDocument65 pagesNervous System and Coordination of Nervous and Endocrine System To Achieve HomeostasisJoseph yvannNo ratings yet
- How Hormones Govern Body ActivitiesDocument4 pagesHow Hormones Govern Body ActivitiesMarvin MelisNo ratings yet
- Esson 3 Feedback Mechanism and Homeostasis StudentDocument27 pagesEsson 3 Feedback Mechanism and Homeostasis StudentJean Marc ReyNo ratings yet
- Topic 13 John Paul Magtalas G BioDocument39 pagesTopic 13 John Paul Magtalas G BioRaon, Simone AndreiNo ratings yet
- NegOr Q4 GenBio2 SLKWeek4 v2Document20 pagesNegOr Q4 GenBio2 SLKWeek4 v2Maribel Ruedas RamosNo ratings yet
- Control and Coordination CL XDocument7 pagesControl and Coordination CL XMukul BanoulaNo ratings yet
- AnscieDocument10 pagesAnscieEstephanie BusligNo ratings yet
- MODULE 2 Ansci 1Document31 pagesMODULE 2 Ansci 1Nelgine GepuitNo ratings yet
- Science 10 Handout - Nervous and EndocrineDocument2 pagesScience 10 Handout - Nervous and EndocrineAngelica MartinNo ratings yet
- Activity 1 Animal SciDocument5 pagesActivity 1 Animal SciKylle BedisNo ratings yet
- Anatomy Fuj JobbyDocument15 pagesAnatomy Fuj JobbyObadah AbdulsattarNo ratings yet
- Endocrine System 1 PDFDocument10 pagesEndocrine System 1 PDFDilNo ratings yet
- Control and Coordination - Class XDocument5 pagesControl and Coordination - Class XMonika Mehan89% (19)
- Topic 3 - Nervous Control in Plants and AnimalsDocument5 pagesTopic 3 - Nervous Control in Plants and AnimalsYara Jeah Ingay BaliliNo ratings yet
- ABPG1103 - Topic 2 - Biological Psychology - 222Document31 pagesABPG1103 - Topic 2 - Biological Psychology - 222nureryani bathowiNo ratings yet
- Q3 - Nervous and Endocrine Systems - Science10 PDFDocument84 pagesQ3 - Nervous and Endocrine Systems - Science10 PDFGabriel Pasumbal100% (1)
- TVBS Nervous SystemDocument20 pagesTVBS Nervous SystemAngel Mae Masa FloresNo ratings yet
- Animal Organ Systems and Homeostasis PDFDocument13 pagesAnimal Organ Systems and Homeostasis PDFAnonymous HXLczq3No ratings yet
- Chapter 3Document109 pagesChapter 3amala raviNo ratings yet
- The Receiving Mechanism: Sense Organs Are Sensitive Nerve Endings Located in Certain Body Parts. They Are Receptors ofDocument5 pagesThe Receiving Mechanism: Sense Organs Are Sensitive Nerve Endings Located in Certain Body Parts. They Are Receptors ofArlene Cerdeña SalcedaNo ratings yet
- Science 10 Quarter 3 Module 3Document15 pagesScience 10 Quarter 3 Module 3Zahra Alexä DavidsonNo ratings yet
- B Science 10 Quarter 3 Module 2 (Week 3)Document17 pagesB Science 10 Quarter 3 Module 2 (Week 3)Daisy Soriano PrestozaNo ratings yet
- NCERT Summary Control & CoordinationDocument12 pagesNCERT Summary Control & Coordinationpadhako.harshNo ratings yet
- Chapter 1 - The Science of Anatomy and Physiology UDocument5 pagesChapter 1 - The Science of Anatomy and Physiology UJess SommervilleNo ratings yet
- Essay B. InggrisDocument9 pagesEssay B. InggrisM.HalilNo ratings yet
- Chapter 7 Bio Class 10Document15 pagesChapter 7 Bio Class 10Aysha KhanNo ratings yet
- Chemical and Nervous SystemDocument42 pagesChemical and Nervous SystemMa Divina Kristi DiscarNo ratings yet
- Psychology PresentationDocument16 pagesPsychology PresentationFasih RehmanNo ratings yet
- Control and Coordination Do You Know Which Organs Make Up The Nervous System?Document22 pagesControl and Coordination Do You Know Which Organs Make Up The Nervous System?So TemptedNo ratings yet
- Class 10 Chapter 7 NotesDocument7 pagesClass 10 Chapter 7 NotesIra LaratteNo ratings yet
- Nervous SystemDocument4 pagesNervous SystemTherese MaleriadoNo ratings yet
- What Is Human Endocrine SystemDocument38 pagesWhat Is Human Endocrine SystemNatukunda DianahNo ratings yet
- Control Coordination NotesDocument6 pagesControl Coordination NotesRaman ChaudharyNo ratings yet
- Discuss and Elaborate The Relationship of The Different Systems Found in The Animal BodyDocument3 pagesDiscuss and Elaborate The Relationship of The Different Systems Found in The Animal BodyJenny V. BajoNo ratings yet
- Cbse Class 10 Science Notes Chapter 7 Control and CoordinationDocument8 pagesCbse Class 10 Science Notes Chapter 7 Control and CoordinationFatema EshrakNo ratings yet
- Control and CoordinationDocument8 pagesControl and CoordinationkkNo ratings yet
- Unit 4 Perception and Coordination 1Document84 pagesUnit 4 Perception and Coordination 1Iker anubisNo ratings yet
- Control and CoordinationDocument11 pagesControl and CoordinationJit AggNo ratings yet
- Control and Coordination: - Prepared by-R.S.Chandanshiv, Principal, JNV, N.C.Hills (Assam)Document30 pagesControl and Coordination: - Prepared by-R.S.Chandanshiv, Principal, JNV, N.C.Hills (Assam)Sheetal Kendre DagadkhairNo ratings yet
- Fall 2022 - ZOO403 - 1 - BC190408115Document4 pagesFall 2022 - ZOO403 - 1 - BC190408115Misbah NoreenNo ratings yet
- Copy-REGULATORY SYSTEM HODocument8 pagesCopy-REGULATORY SYSTEM HORio FirmansyahNo ratings yet
- Week 19 HSBDocument15 pagesWeek 19 HSBedhanassarNo ratings yet
- Form 4 Science Chapter 2Document13 pagesForm 4 Science Chapter 2Cheah Foo Kit50% (2)
- Anatomy and Physiology ReviewerDocument10 pagesAnatomy and Physiology ReviewerMae Christelle FigueroaNo ratings yet
- Preparation in Psychology: Bicol University College of Engineering East Campus LegazpiDocument8 pagesPreparation in Psychology: Bicol University College of Engineering East Campus LegazpiKielNo ratings yet
- Anatomy of The BrainDocument38 pagesAnatomy of The BrainnayanaNo ratings yet
- Departemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraDocument21 pagesDepartemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraFira TasyaNo ratings yet
- Anatomy and Fisiology NoteDocument3 pagesAnatomy and Fisiology NoteIssac LauNo ratings yet
- NScELEC3 WEEK 9 LESSONDocument9 pagesNScELEC3 WEEK 9 LESSONAlyssa Jane AbellonNo ratings yet
- A. MedStu - CNSDocument20 pagesA. MedStu - CNSNwraz AlsahibNo ratings yet
- Nervous System FunctionsDocument10 pagesNervous System FunctionsJovi Leo PunoNo ratings yet
- Departemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraDocument20 pagesDepartemen Fisiologi Fakultas Kedokteran Universitas Sumatera UtaraabbyNo ratings yet
- Gale Researcher Guide for: Overview of Physiology and NeuropsychologyFrom EverandGale Researcher Guide for: Overview of Physiology and NeuropsychologyNo ratings yet
- Intro To Human Anatomy & Physiology: Quick Review Notes Chapter 1From EverandIntro To Human Anatomy & Physiology: Quick Review Notes Chapter 1No ratings yet
- Chapter 2 UNIT IVDocument24 pagesChapter 2 UNIT IVAyen LatosaNo ratings yet
- Chapter 2 UNIT IIDocument10 pagesChapter 2 UNIT IIAyen LatosaNo ratings yet
- Chapter 2 UNIT 1Document19 pagesChapter 2 UNIT 1Ayen LatosaNo ratings yet
- Cardiovascular System AnatomyDocument54 pagesCardiovascular System AnatomyAyen LatosaNo ratings yet
- Blood Pressure - Blood Pressure ChartDocument3 pagesBlood Pressure - Blood Pressure ChartMahmoud MaherNo ratings yet
- Activity 8 - Blood and HematopoiesisDocument8 pagesActivity 8 - Blood and HematopoiesisMarissa CordovaNo ratings yet
- Ex 5 Activity 3Document4 pagesEx 5 Activity 3Rafeik05100% (2)
- Name - Soni Ritika Rajesh, Group - GM20-054-2A Physiology Week 03 TasksDocument10 pagesName - Soni Ritika Rajesh, Group - GM20-054-2A Physiology Week 03 TasksRitika SoniNo ratings yet
- Hemophilia: Joanna Marie de Guzman RNDocument35 pagesHemophilia: Joanna Marie de Guzman RNJoan VenidaNo ratings yet
- Ineffective Tissue PerfussionDocument2 pagesIneffective Tissue Perfussionmonico39No ratings yet
- Blood Groups SystemsDocument7 pagesBlood Groups SystemsMir jaliesNo ratings yet
- 【Zybio】Z3 Service Training PPT-Distributor 20200110Document116 pages【Zybio】Z3 Service Training PPT-Distributor 20200110Fermin Tafur Lliuya100% (1)
- Disciplina AquasDocument84 pagesDisciplina AquasMoises TellezNo ratings yet
- 7A Cells, Tissues and Organs Booklet - Doc-1Document16 pages7A Cells, Tissues and Organs Booklet - Doc-1Majdoline SadeddineNo ratings yet
- Neonatal Resuscitation PDFDocument1 pageNeonatal Resuscitation PDFOdessa EnriquezNo ratings yet
- Management and Prognosis of The Thalassemias - UpToDateDocument45 pagesManagement and Prognosis of The Thalassemias - UpToDatesushi37No ratings yet
- 1 Hema LecDocument5 pages1 Hema LecAngela ReyesNo ratings yet
- Basic Science JSS2 Question BankDocument3 pagesBasic Science JSS2 Question BankIsaac100% (1)
- Cardiogenic Shock NCLEX Review QuizDocument6 pagesCardiogenic Shock NCLEX Review QuizRegie Marie EvangelistaNo ratings yet
- Exploring Science Edition © Pearson Education Limited 2008Document2 pagesExploring Science Edition © Pearson Education Limited 2008Victor Barber Sanchis100% (1)
- Blood Banking Course BookDocument2 pagesBlood Banking Course BookShukr Wesman BlbasNo ratings yet
- Natural Eye CareDocument222 pagesNatural Eye Careskribdot100% (3)
- The Effects of Lifestyle To Respiratory and Circulatory SystemsDocument11 pagesThe Effects of Lifestyle To Respiratory and Circulatory SystemsSharalyn Sia100% (1)
- Circulatory and Respiratory System, Working TogetherDocument32 pagesCirculatory and Respiratory System, Working TogetherRhona AngelaNo ratings yet
- S4 Mock 1 2006 Biology P1Document8 pagesS4 Mock 1 2006 Biology P1cyber sec100% (1)
- Ha&P - Semi ReviewerDocument71 pagesHa&P - Semi ReviewerLittle StuartNo ratings yet
- Vox Sanguin Maret 2022 - OptDocument192 pagesVox Sanguin Maret 2022 - Optrsdarsono labNo ratings yet
- Kaigo Prometric Lesson (Eng)Document108 pagesKaigo Prometric Lesson (Eng)Ysmech SalazarNo ratings yet
- Iron MetabolismDocument3 pagesIron MetabolismlucaNo ratings yet
- Isaac Typhoid NegativeDocument5 pagesIsaac Typhoid Negativeganesh khondeNo ratings yet
- Blood Gas HandbookDocument112 pagesBlood Gas HandbookNuno Silva100% (1)
- Hkcee Biology - 4.6 Transport in Humans - P.1Document13 pagesHkcee Biology - 4.6 Transport in Humans - P.1irisyyy27No ratings yet