
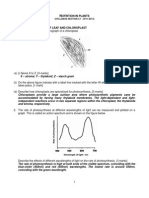
Uncoupling To Survive
Uncoupling To Survive
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- PHOTOSYNTHESISDocument17 pagesPHOTOSYNTHESISDave Degaetano100% (3)
- Gene MiningDocument44 pagesGene MiningVIKAS DOGRA100% (10)
- Gel Electrophoresis ProjectDocument3 pagesGel Electrophoresis Projectvidhya sureshNo ratings yet
- Models of Dna ReplicationDocument11 pagesModels of Dna ReplicationtoshiNo ratings yet
- What Makes Animal Cells DifferentDocument14 pagesWhat Makes Animal Cells DifferentGian Carlo Angon100% (1)
- DNA LibraryDocument73 pagesDNA LibraryDGA GAMINGNo ratings yet
- General Biology 1 Module 8 Q1Document21 pagesGeneral Biology 1 Module 8 Q1Pril Gueta100% (4)
- Receptors: Types Structure FunctionsDocument44 pagesReceptors: Types Structure Functionsmaham1085No ratings yet
- DR - Sc.nahida SrabovićDocument70 pagesDR - Sc.nahida SrabovićMr. LenardNo ratings yet
- Ligation Sequencing gDNA V14 - Whole Genome Amplification (SQK-LSK114) - MinionDocument8 pagesLigation Sequencing gDNA V14 - Whole Genome Amplification (SQK-LSK114) - MinionDevansh PanwarNo ratings yet
- 2022 05 Benchmarking Transcriptomics Methods Transcript CellDocument3 pages2022 05 Benchmarking Transcriptomics Methods Transcript CellTrần Viết Nhân NghịNo ratings yet
- January 2009 QP - Unit 1 Edexcel Biology A-LevelDocument24 pagesJanuary 2009 QP - Unit 1 Edexcel Biology A-LevelFatma ZorluNo ratings yet
- Chemistry Is The Logic of Biological Phenomena: To Accompany Biochemistry, 2/e by Reginald Garrett and Charles GrishamDocument32 pagesChemistry Is The Logic of Biological Phenomena: To Accompany Biochemistry, 2/e by Reginald Garrett and Charles GrishamTarun AroraNo ratings yet
- Biol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairDocument6 pagesBiol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairJohn Paradise100% (1)
- Review - Milk Protein, Types and Structure, Synthesis of Casein ResearchgateDocument22 pagesReview - Milk Protein, Types and Structure, Synthesis of Casein ResearchgateNaddiaNo ratings yet
- Amersham Histraphp InstructDocument20 pagesAmersham Histraphp InstructkostrenarijekaNo ratings yet
- Microbiology Chapter 3 SlidesDocument21 pagesMicrobiology Chapter 3 Slideseileen_muu485No ratings yet
- Lehninger Multiple ChoiceDocument13 pagesLehninger Multiple ChoiceHugo DuarteNo ratings yet
- CHY 47 Topic 4 2nd Sem 2018-2019 Protein Amino Acid PDFDocument129 pagesCHY 47 Topic 4 2nd Sem 2018-2019 Protein Amino Acid PDFHA RENo ratings yet
- Circular Dichroism PDFDocument7 pagesCircular Dichroism PDFprakas.rao39695No ratings yet
- Genetic Polymorphisms Personalized MedicineDocument38 pagesGenetic Polymorphisms Personalized MedicineBayu WisnuNo ratings yet
- Monod Nobel LectureDocument22 pagesMonod Nobel LectureAnaNo ratings yet
- Ecor1 (0/4363) : 18 April 2023Document3 pagesEcor1 (0/4363) : 18 April 2023Thandeka NseleNo ratings yet
- Cellular Respiration GlycolysisDocument1 pageCellular Respiration GlycolysisReigne CuevasNo ratings yet
- Serodos Plus Valores Lot 0004Document3 pagesSerodos Plus Valores Lot 0004luisoft88No ratings yet
- Example of Explanation TextDocument1 pageExample of Explanation TextEko Pamungkas50% (2)
- Cell Division ReviewDocument7 pagesCell Division ReviewBLlahniesss Love Ken100% (1)
- Review of Cell Division BSC StudentsDocument64 pagesReview of Cell Division BSC StudentsKripa SusanNo ratings yet
- Size Exclusion ChortamographyDocument124 pagesSize Exclusion ChortamographyViviana CastilloNo ratings yet




































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- PHOTOSYNTHESISDocument17 pagesPHOTOSYNTHESISDave Degaetano100% (3)
- Gene MiningDocument44 pagesGene MiningVIKAS DOGRA100% (10)
- Gel Electrophoresis ProjectDocument3 pagesGel Electrophoresis Projectvidhya sureshNo ratings yet
- Models of Dna ReplicationDocument11 pagesModels of Dna ReplicationtoshiNo ratings yet
- What Makes Animal Cells DifferentDocument14 pagesWhat Makes Animal Cells DifferentGian Carlo Angon100% (1)
- DNA LibraryDocument73 pagesDNA LibraryDGA GAMINGNo ratings yet
- General Biology 1 Module 8 Q1Document21 pagesGeneral Biology 1 Module 8 Q1Pril Gueta100% (4)
- Receptors: Types Structure FunctionsDocument44 pagesReceptors: Types Structure Functionsmaham1085No ratings yet
- DR - Sc.nahida SrabovićDocument70 pagesDR - Sc.nahida SrabovićMr. LenardNo ratings yet
- Ligation Sequencing gDNA V14 - Whole Genome Amplification (SQK-LSK114) - MinionDocument8 pagesLigation Sequencing gDNA V14 - Whole Genome Amplification (SQK-LSK114) - MinionDevansh PanwarNo ratings yet
- 2022 05 Benchmarking Transcriptomics Methods Transcript CellDocument3 pages2022 05 Benchmarking Transcriptomics Methods Transcript CellTrần Viết Nhân NghịNo ratings yet
- January 2009 QP - Unit 1 Edexcel Biology A-LevelDocument24 pagesJanuary 2009 QP - Unit 1 Edexcel Biology A-LevelFatma ZorluNo ratings yet
- Chemistry Is The Logic of Biological Phenomena: To Accompany Biochemistry, 2/e by Reginald Garrett and Charles GrishamDocument32 pagesChemistry Is The Logic of Biological Phenomena: To Accompany Biochemistry, 2/e by Reginald Garrett and Charles GrishamTarun AroraNo ratings yet
- Biol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairDocument6 pagesBiol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairJohn Paradise100% (1)
- Review - Milk Protein, Types and Structure, Synthesis of Casein ResearchgateDocument22 pagesReview - Milk Protein, Types and Structure, Synthesis of Casein ResearchgateNaddiaNo ratings yet
- Amersham Histraphp InstructDocument20 pagesAmersham Histraphp InstructkostrenarijekaNo ratings yet
- Microbiology Chapter 3 SlidesDocument21 pagesMicrobiology Chapter 3 Slideseileen_muu485No ratings yet
- Lehninger Multiple ChoiceDocument13 pagesLehninger Multiple ChoiceHugo DuarteNo ratings yet
- CHY 47 Topic 4 2nd Sem 2018-2019 Protein Amino Acid PDFDocument129 pagesCHY 47 Topic 4 2nd Sem 2018-2019 Protein Amino Acid PDFHA RENo ratings yet
- Circular Dichroism PDFDocument7 pagesCircular Dichroism PDFprakas.rao39695No ratings yet
- Genetic Polymorphisms Personalized MedicineDocument38 pagesGenetic Polymorphisms Personalized MedicineBayu WisnuNo ratings yet
- Monod Nobel LectureDocument22 pagesMonod Nobel LectureAnaNo ratings yet
- Ecor1 (0/4363) : 18 April 2023Document3 pagesEcor1 (0/4363) : 18 April 2023Thandeka NseleNo ratings yet
- Cellular Respiration GlycolysisDocument1 pageCellular Respiration GlycolysisReigne CuevasNo ratings yet
- Serodos Plus Valores Lot 0004Document3 pagesSerodos Plus Valores Lot 0004luisoft88No ratings yet
- Example of Explanation TextDocument1 pageExample of Explanation TextEko Pamungkas50% (2)
- Cell Division ReviewDocument7 pagesCell Division ReviewBLlahniesss Love Ken100% (1)
- Review of Cell Division BSC StudentsDocument64 pagesReview of Cell Division BSC StudentsKripa SusanNo ratings yet
- Size Exclusion ChortamographyDocument124 pagesSize Exclusion ChortamographyViviana CastilloNo ratings yet