Download as pdf or txt
You might also like
- Internship ReportDocument11 pagesInternship ReportgeorgeNo ratings yet
- Role of Progesterone in Tlr4-Myd88-Dependent Signaling Pathway in Pre-EclampsiaDocument5 pagesRole of Progesterone in Tlr4-Myd88-Dependent Signaling Pathway in Pre-EclampsiaCarl Enrique PSNo ratings yet
- tmpD776 TMPDocument6 pagestmpD776 TMPFrontiersNo ratings yet
- Tumor necrosis factor α stimulates expression and secretion of urokinase plasminogen activator in human dental pulp cellsDocument8 pagesTumor necrosis factor α stimulates expression and secretion of urokinase plasminogen activator in human dental pulp cellsNadya PurwantyNo ratings yet
- Macrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin ReceptorsDocument8 pagesMacrophage-Stimulating Peptides VKGFY and Cyclo (VKGFY) Act Through Nonopioid B-Endorphin Receptorsvanessa_murillo_5No ratings yet
- 39 809Document7 pages39 809windyndaNo ratings yet
- MMR 2 5 857 PDFDocument8 pagesMMR 2 5 857 PDFguugle gogleNo ratings yet
- Leptin 1Document9 pagesLeptin 1vestibuleNo ratings yet
- A New Cell-Based Assay To Evaluate Myogenesis in Mouse MyoblastDocument11 pagesA New Cell-Based Assay To Evaluate Myogenesis in Mouse MyoblastWendy Lilian Gómez MonsivaisNo ratings yet
- (Kyung Hee, Et Al, 2006) PGDocument4 pages(Kyung Hee, Et Al, 2006) PGWinda AlpiniawatiNo ratings yet
- Piperlongumine Attenuates Il1induced Inflammatory Response in ChondrocytesDocument6 pagesPiperlongumine Attenuates Il1induced Inflammatory Response in ChondrocytesRiya CassendraNo ratings yet
- Ertao 2016Document7 pagesErtao 2016chemistpl420No ratings yet
- Promotion by Collagen Tripeptide of Type I Collagen Gene Expression in Human Osteoblastic Cells and Fracture Healing of Rat Femur (骨折修復) -已解鎖Document8 pagesPromotion by Collagen Tripeptide of Type I Collagen Gene Expression in Human Osteoblastic Cells and Fracture Healing of Rat Femur (骨折修復) -已解鎖Sveto TasevNo ratings yet
- Antikanker 3Document5 pagesAntikanker 3Namira NaaziahNo ratings yet
- (17417899 - Reproduction) The Human Corpus Luteum - Which Cells Have Progesterone ReceptorsDocument9 pages(17417899 - Reproduction) The Human Corpus Luteum - Which Cells Have Progesterone Receptorsتقوى اللهNo ratings yet
- 13 151 2Document8 pages13 151 2B-win IrawanNo ratings yet
- Jcinvest00043 0362Document8 pagesJcinvest00043 0362api-26848049No ratings yet
- Journal 6Document7 pagesJournal 6Claudia JessicaNo ratings yet
- Mco 12 06 0533Document8 pagesMco 12 06 0533Anonymous Wcj4C3jNo ratings yet
- SJ BJP 0706338Document8 pagesSJ BJP 0706338Rogério HitsugayaNo ratings yet
- Cao 2015Document10 pagesCao 2015joselouisngNo ratings yet
- tmp2977 TMPDocument7 pagestmp2977 TMPFrontiersNo ratings yet
- Effet Glucane Reishi 19 Janv 2023Document8 pagesEffet Glucane Reishi 19 Janv 2023ilyNo ratings yet
- A Small Ras-Like Protein Ray-Rab1c Modulates The P53-Regulating Activity of PRPKDocument9 pagesA Small Ras-Like Protein Ray-Rab1c Modulates The P53-Regulating Activity of PRPKCarlos Martinez SalgadoNo ratings yet
- Molecular Analysis of The Pathophysiological Binding of The Platelet Aggregation-Inducing Factor Podoplanin To The C-Type Lectin-Like Receptor CLEC-2Document8 pagesMolecular Analysis of The Pathophysiological Binding of The Platelet Aggregation-Inducing Factor Podoplanin To The C-Type Lectin-Like Receptor CLEC-2zsyedphd1No ratings yet
- Bis N NorgliovictinDocument12 pagesBis N NorgliovictinMichael RigonNo ratings yet
- Projet Ruxo Nikola AnglaisDocument12 pagesProjet Ruxo Nikola AnglaisNikola ૐ Mantov ஓம்No ratings yet
- NRP 9 E18Document6 pagesNRP 9 E18wawansiswadi01No ratings yet
- Expression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedDocument0 pagesExpression of A Gene Encoding Trypanosoma Congolense Putative Abc1 Family Protein Is Developmentally RegulatedMaísa FonsecaNo ratings yet
- Emdogain-Regulated Gene Expression in Palatal Fibroblasts Requires TGF-bRI Kinase SignalingDocument10 pagesEmdogain-Regulated Gene Expression in Palatal Fibroblasts Requires TGF-bRI Kinase SignalingMaria-Lavinia HoinaruNo ratings yet
- Cisplatin Cell Cicle ArrestDocument7 pagesCisplatin Cell Cicle ArrestGabriele KrauseNo ratings yet
- CPG Intranasal en La Prevencion de MetastasisDocument7 pagesCPG Intranasal en La Prevencion de MetastasisRamiro J. Rodriguez GarciaNo ratings yet
- Thymol Inhibits LPS-Stimulated In flammatory Response via Down-Regulation of NF-κB and MAPK Signaling Pathways in Mouse Mammary Epithelial CellsDocument9 pagesThymol Inhibits LPS-Stimulated In flammatory Response via Down-Regulation of NF-κB and MAPK Signaling Pathways in Mouse Mammary Epithelial CellsGianluca FrancoNo ratings yet
- Environmental Mutagen Society of Japan 19th AnnualDocument2 pagesEnvironmental Mutagen Society of Japan 19th AnnualQ-MT Branch HQ 497 OM&MNo ratings yet
- 2000-Drug Development ResearchDocument10 pages2000-Drug Development ResearchEmerson CasaliNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument6 pagesBiochemical and Biophysical Research CommunicationsSergeat18BNo ratings yet
- Fluoxtine InducedDocument7 pagesFluoxtine InducedsufaNo ratings yet
- LeptocarpinaDocument7 pagesLeptocarpinaJ Nicolas MJNo ratings yet
- Helicobacter Pylori: Upregulates Prion Protein Expression in Gastric Mucosa: A Possible Link To Prion DiseaseDocument6 pagesHelicobacter Pylori: Upregulates Prion Protein Expression in Gastric Mucosa: A Possible Link To Prion DiseaseNadeem MalikNo ratings yet
- Ajpendo 90369 2008Document9 pagesAjpendo 90369 2008Maria Virgina Perdomo De Los SantosNo ratings yet
- Dietary L-Arginine Supplementation Reduces Methotrexate Induced Intestinal Mucosal Injury in RatDocument9 pagesDietary L-Arginine Supplementation Reduces Methotrexate Induced Intestinal Mucosal Injury in RatMohamed IsmailNo ratings yet
- Osteopontin is essential for IL‐1β production and apoptosis in peri‐implantitisDocument9 pagesOsteopontin is essential for IL‐1β production and apoptosis in peri‐implantitisBagis Emre GulNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument6 pagesBiochemical and Biophysical Research CommunicationsSebastián GallegosNo ratings yet
- Seong 2015Document7 pagesSeong 2015Jocilene Dantas Torres NascimentoNo ratings yet
- Phosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersDocument11 pagesPhosphatidylinositol 3 - Kinase Signalling Supports Cell Height in Established Epithelial MonolayersMohammed Elias AlamNo ratings yet
- Effect of Low Dose Quercetin and CancerDocument21 pagesEffect of Low Dose Quercetin and CancerMetcher Maa AkhiNo ratings yet
- Curcumin - ECMDocument7 pagesCurcumin - ECMBABANG GAGAHNo ratings yet
- Journal CancerDocument17 pagesJournal CancerbeklogNo ratings yet
- MMP10 NSCLCDocument9 pagesMMP10 NSCLCbhaskarNo ratings yet
- Refolding and Characterization of RecombDocument6 pagesRefolding and Characterization of RecombMobarakNo ratings yet
- Shen 2018Document6 pagesShen 2018Arturo JiménezNo ratings yet
- Epigallocatechin-3-Gallate Suppresses TNF-of MMP-1 and - 3 in Rheumatoid Arthritis Synovial WbroblastsDocument7 pagesEpigallocatechin-3-Gallate Suppresses TNF-of MMP-1 and - 3 in Rheumatoid Arthritis Synovial WbroblastsTharuka ChandrasiriNo ratings yet
- Pi Is 0021925819770471Document6 pagesPi Is 0021925819770471ivette ramirezNo ratings yet
- Ahmed 2006Document9 pagesAhmed 2006Tharuka ChandrasiriNo ratings yet
- Delfinidina PaperDocument13 pagesDelfinidina PaperCristobal ParraNo ratings yet
- Poster-Presentaion-Abstracts ASPIRE2017Document241 pagesPoster-Presentaion-Abstracts ASPIRE2017HAVIZ YUADNo ratings yet
- Clinacanthus Nutans ExtractsDocument11 pagesClinacanthus Nutans ExtractsNop PiromNo ratings yet
- Corea Del Sur 2019 SODIUMGLUCOSE COTRANSPORTER 2 INHIBOTOR FORDocument1 pageCorea Del Sur 2019 SODIUMGLUCOSE COTRANSPORTER 2 INHIBOTOR FORLaura GarciaNo ratings yet
- Ijmm 25 4 593 PDFDocument8 pagesIjmm 25 4 593 PDFNickNo ratings yet
- Transplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Document128 pagesTransplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Enrique Moreno GonzálezNo ratings yet
- Lilbm3: Adita Ayu Aprilia 30101407112Document16 pagesLilbm3: Adita Ayu Aprilia 30101407112Adita AyuNo ratings yet
- EASD2020 FinalProgrammeDocument318 pagesEASD2020 FinalProgrammeJia-PeiWuNo ratings yet
- Evsu - Bsed-Biosci - Anatomy and Physiology Study GuideDocument3 pagesEvsu - Bsed-Biosci - Anatomy and Physiology Study GuideMARY ANGELA LERIOSNo ratings yet
- Understanding Cholesterol: Classification of A LipoproteinDocument16 pagesUnderstanding Cholesterol: Classification of A LipoproteinJacky FaragNo ratings yet
- Observation of The Patient When They Walk in - : HistoryDocument19 pagesObservation of The Patient When They Walk in - : Historyajaykumar soddalaNo ratings yet
- Disorders of Cardiac RhythmDocument66 pagesDisorders of Cardiac RhythmslyfoxkittyNo ratings yet
- Erik Van Woensel Classical Homeopathy Evidence Based Medicine Vol. 2Document23 pagesErik Van Woensel Classical Homeopathy Evidence Based Medicine Vol. 2Sohail LatifNo ratings yet
- IP Address: 157.47.128.69 Date: 06/09/2020 Time: 14:06:43 PMDocument2 pagesIP Address: 157.47.128.69 Date: 06/09/2020 Time: 14:06:43 PMSanju YadavNo ratings yet
- Read online textbook A Compendium Of Neuropsychological Tests Fundamentals Of Neuropsychological Assessment And Test Reviews For Clinical Practice 4E Sep 21 2021_0199856184_Oxford University Press 4Th Edition Sh ebook all chapter pdfDocument23 pagesRead online textbook A Compendium Of Neuropsychological Tests Fundamentals Of Neuropsychological Assessment And Test Reviews For Clinical Practice 4E Sep 21 2021_0199856184_Oxford University Press 4Th Edition Sh ebook all chapter pdflisa.roper448100% (9)
- Inhalational AgentsDocument12 pagesInhalational Agentsdrhiwaomer100% (1)
- Biocontrol Activity of Bacillus Against A GFP-marked Pseudomonas Syringae Pv. Tomato On Tomato PhylloplaneDocument11 pagesBiocontrol Activity of Bacillus Against A GFP-marked Pseudomonas Syringae Pv. Tomato On Tomato PhylloplaneRoberto Lanna FilhoNo ratings yet
- Digestive SystemDocument29 pagesDigestive Systemsugar kookie0% (1)
- Drop Hallux PDFDocument5 pagesDrop Hallux PDFFiras BerroNo ratings yet
- Aromatherapy Massage For Neuro PDFDocument11 pagesAromatherapy Massage For Neuro PDFTitin JuherniNo ratings yet
- Holistic Approach To HealthDocument30 pagesHolistic Approach To Healthmaeydel0% (1)
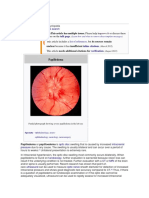
- Papilledema: Jump To Navigationjump To SearchDocument4 pagesPapilledema: Jump To Navigationjump To SearchEri DooNo ratings yet
- The Impact of Physical Activities On The Health of AdultsDocument12 pagesThe Impact of Physical Activities On The Health of AdultsJUNAID AHMEDNo ratings yet
- Annotated BibliographyDocument3 pagesAnnotated Bibliographyapi-316836226No ratings yet
- Hepatitis B Surface AntigenDocument5 pagesHepatitis B Surface AntigenTantri D'des KriegersNo ratings yet
- Knowledge and Attitude Regarding Cervical Cancer Vaccination Among Female Undergraduates at Kampala International University-Western CampusDocument9 pagesKnowledge and Attitude Regarding Cervical Cancer Vaccination Among Female Undergraduates at Kampala International University-Western CampusKIU PUBLICATION AND EXTENSIONNo ratings yet
- Kondisi Pencemaran Lingkungan Berdasarkan Parameter Mikrobiologis Di Sekitar Muara Sungai Cimandiri, Teluk Pelabuhan Ratu, Jawa BaratDocument6 pagesKondisi Pencemaran Lingkungan Berdasarkan Parameter Mikrobiologis Di Sekitar Muara Sungai Cimandiri, Teluk Pelabuhan Ratu, Jawa BaratNurulNo ratings yet
- Multiple Choice Questions: Control of Body MovementDocument20 pagesMultiple Choice Questions: Control of Body MovementwanderagroNo ratings yet
- Ranch Management Plan-Darien Holdings PDFDocument12 pagesRanch Management Plan-Darien Holdings PDFKealeboga Duece Thobolo100% (1)
- STELLAR Study (Rosuvastatina)Document44 pagesSTELLAR Study (Rosuvastatina)ijido_sc100% (1)
- Terebinthiniae Oleum: Other Names: Made FromDocument30 pagesTerebinthiniae Oleum: Other Names: Made FromDohn Joe100% (1)
- Radiology of GITDocument12 pagesRadiology of GITAustine OsaweNo ratings yet
- Plasma Banks: Our New Knight in Shining Armour?Document2 pagesPlasma Banks: Our New Knight in Shining Armour?Mya StoneNo ratings yet
- Traditional Herbal Drugs of Southern Uganda, II - Literature Analysis and Antimicrobial AssaysDocument22 pagesTraditional Herbal Drugs of Southern Uganda, II - Literature Analysis and Antimicrobial AssaysMinyty LeNo ratings yet
- The Cell - PinedaDocument13 pagesThe Cell - PinedaOHMYK33NZYNo ratings yet