Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- SadlerDocument1 pageSadlerryan aniceteNo ratings yet
- SME VOL2 - Auto - x2Document1 pageSME VOL2 - Auto - x2ryan aniceteNo ratings yet
- SME VOL1 - Auto - x2Document1 pageSME VOL1 - Auto - x2ryan aniceteNo ratings yet
- Pipe Thread EngagementDocument2 pagesPipe Thread Engagementryan aniceteNo ratings yet
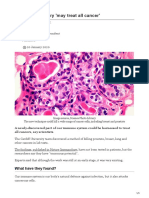
- Immune Discovery May Treat All Cancer - 2020jan20Document5 pagesImmune Discovery May Treat All Cancer - 2020jan20ryan aniceteNo ratings yet
- Bisphenol A Elimination by TAML+Hydrogen Peroxide - Collins 2017Document29 pagesBisphenol A Elimination by TAML+Hydrogen Peroxide - Collins 2017ryan aniceteNo ratings yet
- Qty of Joints For FullweldingDocument1 pageQty of Joints For Fullweldingryan aniceteNo ratings yet
- CA A Cancer J Clinicians - July August 1995 - Hortobagyi - Current Status of Adjuvant Systemic Therapy For Primary BreastDocument28 pagesCA A Cancer J Clinicians - July August 1995 - Hortobagyi - Current Status of Adjuvant Systemic Therapy For Primary Breastryan aniceteNo ratings yet
- An Unconventional Weapon Against Cancer - 2020may02Document3 pagesAn Unconventional Weapon Against Cancer - 2020may02ryan aniceteNo ratings yet
- On The Design and Construction of AlgebrDocument11 pagesOn The Design and Construction of Algebrryan aniceteNo ratings yet
- JACK A Process Algebra Implementation inDocument200 pagesJACK A Process Algebra Implementation inryan aniceteNo ratings yet
- How To Make AXIOM Into A ScratchpadDocument10 pagesHow To Make AXIOM Into A Scratchpadryan aniceteNo ratings yet
- PVC Clear Schedule 80Document1 pagePVC Clear Schedule 80ryan aniceteNo ratings yet
- Autocad 2D: Training ManualDocument94 pagesAutocad 2D: Training Manualryan aniceteNo ratings yet
- 1.3 Schedule: Schedule Work Performed This PeriodDocument1 page1.3 Schedule: Schedule Work Performed This Periodryan aniceteNo ratings yet
- Symbolic Script Programming For Java: June 2009Document17 pagesSymbolic Script Programming For Java: June 2009ryan aniceteNo ratings yet
- Interfacing R E D U C E To Java: and andDocument2 pagesInterfacing R E D U C E To Java: and andryan aniceteNo ratings yet
- Joses2 FinalDocument8 pagesJoses2 Finalryan aniceteNo ratings yet
- Excel Vba-Based Solution To Pipe Flow Measurement Problem: Spreadsheets in Education (Ejsie)Document16 pagesExcel Vba-Based Solution To Pipe Flow Measurement Problem: Spreadsheets in Education (Ejsie)ryan aniceteNo ratings yet
- Design of A Digital Mathematical Library For Science Technology and Education - April 1999Document14 pagesDesign of A Digital Mathematical Library For Science Technology and Education - April 1999ryan aniceteNo ratings yet
- Installation Manual: System Air ConditionersDocument23 pagesInstallation Manual: System Air Conditionersryan aniceteNo ratings yet
- Inside Reduce: Arthur C. Norman Raffaele VitoloDocument82 pagesInside Reduce: Arthur C. Norman Raffaele Vitoloryan aniceteNo ratings yet
- FXZQ-MVE DatabookDocument22 pagesFXZQ-MVE Databookryan aniceteNo ratings yet