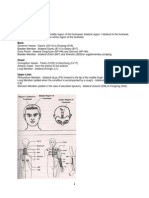
Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5825)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (903)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- 1.atlas Hematologi Diggs PDFDocument128 pages1.atlas Hematologi Diggs PDFPuspa YaNi100% (6)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Testicular BreathingDocument8 pagesTesticular BreathingYCY57% (30)
- Wildlife Fact File - Birds - 61-70Document20 pagesWildlife Fact File - Birds - 61-70ClearMind84100% (1)
- Witter 2011Document12 pagesWitter 2011Hector UsamanNo ratings yet
- Mcmahon 1989Document4 pagesMcmahon 1989Hector UsamanNo ratings yet
- 2392 7124 1 PBDocument4 pages2392 7124 1 PBHector UsamanNo ratings yet
- CVBD Ehrlichiosis 190225-Web PDFDocument2 pagesCVBD Ehrlichiosis 190225-Web PDFHector UsamanNo ratings yet
- Summative Task Unit 1Document1 pageSummative Task Unit 1Likisha RaffyNo ratings yet
- Arda2014 PDFDocument11 pagesArda2014 PDFAnonymous bFcaYbRROCNo ratings yet
- Homoeopathy and Polarity AnalysisDocument24 pagesHomoeopathy and Polarity Analysissherdil Khan100% (1)
- Exercicio Aula 5Document3 pagesExercicio Aula 5Felipe MacielNo ratings yet
- COMMUNICATION WorksheetsDocument5 pagesCOMMUNICATION WorksheetsGwyneth DalupingNo ratings yet
- P.3 LITERAY 1 Activity Wk.1Document5 pagesP.3 LITERAY 1 Activity Wk.1RokNo ratings yet
- NCP - Acute PainDocument3 pagesNCP - Acute PainRoxanne NavarroNo ratings yet
- Guasha 50 PatologiiDocument55 pagesGuasha 50 PatologiiDan Alex100% (2)
- Nie YinniangDocument6 pagesNie YinniangJames SmithNo ratings yet
- Introduction To Artificial Neural NetworksDocument73 pagesIntroduction To Artificial Neural NetworksMuhammad Abd JalilNo ratings yet
- Epi Dermi OlogyDocument11 pagesEpi Dermi OlogyBindashboy00% (1)
- Code of Practice CompleteDocument9 pagesCode of Practice CompleteAlkistis MarinakiNo ratings yet
- AnimalsDocument2 pagesAnimalsMichelle� Calista SalimNo ratings yet
- Unit 7 How Tall Are YouDocument4 pagesUnit 7 How Tall Are YouLidyaRNo ratings yet
- Đề trường vòng 1 2019Document8 pagesĐề trường vòng 1 2019Hà AnhNo ratings yet
- D&D 5e The Rising TempestDocument21 pagesD&D 5e The Rising TempestJoão Guilherme assim100% (3)
- Chinese HoroscopeDocument61 pagesChinese HoroscopeKarl Jacobs100% (3)
- Neck MassDocument13 pagesNeck MassY3153NSNo ratings yet
- Testicular Torsion RiuDocument28 pagesTesticular Torsion RiuDwiyanti OktaviaNo ratings yet
- Rhetorical Analysis 1 EssayDocument3 pagesRhetorical Analysis 1 EssaymayhuartNo ratings yet
- Vegan EarthDocument1,001 pagesVegan EarthAnonymous pKuORTo0% (1)
- Importance of Bhujangasana in Daily LifeDocument6 pagesImportance of Bhujangasana in Daily LifeEditor IJTSRDNo ratings yet
- 0510 s18 QP 21Document16 pages0510 s18 QP 21IamEnszNo ratings yet
- Pest of Oil Palm 2010-AprDocument12 pagesPest of Oil Palm 2010-AprDhianRhanyPratiwiNo ratings yet
- Live Fish List 6 8 21Document17 pagesLive Fish List 6 8 21Jane MoselleNo ratings yet
- 20 Questions About Myself: Answer The Following QuestionsDocument41 pages20 Questions About Myself: Answer The Following QuestionsKure RukminiNo ratings yet
- Ilovepdf - Merged (3) - 131-140Document10 pagesIlovepdf - Merged (3) - 131-140kanishka saxenaNo ratings yet