
Himmelsbach 2002
Himmelsbach 2002
You might also like
- Balance Sheet & Income Statement For Hospital PDFDocument45 pagesBalance Sheet & Income Statement For Hospital PDFAdrianus PramudhitaNo ratings yet
- Kytococcus Sedentarius The Organism AssociatedDocument7 pagesKytococcus Sedentarius The Organism AssociatedMilena TomásNo ratings yet
- Antibacterial Principles From The Bark of Terminalia Arjuna: Scientific CorrespondenceDocument3 pagesAntibacterial Principles From The Bark of Terminalia Arjuna: Scientific CorrespondenceAshwani GaurNo ratings yet
- 1981 - Ekramoddoullah, Kisil, Sehon - Isolation of Allergenically Active Cytochrome C From Kentucky Blue Grass PollenDocument10 pages1981 - Ekramoddoullah, Kisil, Sehon - Isolation of Allergenically Active Cytochrome C From Kentucky Blue Grass Pollenpond_1993No ratings yet
- Nile Red - Kimura PDFDocument8 pagesNile Red - Kimura PDFSharif M Mizanur RahmanNo ratings yet
- Using Eclipta Prostrata (L.) Against Ht-29 Cell LineDocument5 pagesUsing Eclipta Prostrata (L.) Against Ht-29 Cell LineVardhana JanakiramanNo ratings yet
- Production, Purification and Characterization of An Extracellular Keratinase From Lysobacter NCIMB 9497Document6 pagesProduction, Purification and Characterization of An Extracellular Keratinase From Lysobacter NCIMB 9497Faisal AwanNo ratings yet
- EFRTDocument7 pagesEFRTDiego Fernando Jaimes GonzalezNo ratings yet
- Extraction and Characterization of Cellulose Nanocrystals From Corn StoverDocument7 pagesExtraction and Characterization of Cellulose Nanocrystals From Corn StoverBlessing HarvestNo ratings yet
- Structural Characterization of Cellulose ObtainedDocument5 pagesStructural Characterization of Cellulose ObtainedRezza MarcellaNo ratings yet
- Silva 2013Document4 pagesSilva 2013capau87No ratings yet
- Siedel 1984 Aquacultural-EngineeringDocument14 pagesSiedel 1984 Aquacultural-EngineeringJorge RodriguezNo ratings yet
- Binding and Uptake of Wheat Germ Agglutinin-Grafted PLGA-Nanospheres by Caco-2 MonolayersDocument7 pagesBinding and Uptake of Wheat Germ Agglutinin-Grafted PLGA-Nanospheres by Caco-2 MonolayerszeminkNo ratings yet
- Ravic Hand Ran 2016Document10 pagesRavic Hand Ran 2016Xuân LâmNo ratings yet
- WWW - Cjs 200902 0005Document8 pagesWWW - Cjs 200902 0005Uchiha AbdurrahmanNo ratings yet
- Pääbo Mitochondrial NAR 1988Document14 pagesPääbo Mitochondrial NAR 1988jaafari mohamedNo ratings yet
- Simultaneous Determination of Quinolones in Fish by Liquid Chromatography Coupled With Fluorescence Detection: Comparison of Sub-2 M Particles and Conventional C ColumnsDocument27 pagesSimultaneous Determination of Quinolones in Fish by Liquid Chromatography Coupled With Fluorescence Detection: Comparison of Sub-2 M Particles and Conventional C Columnsnguyen baoNo ratings yet
- Kobanana2 PDFDocument8 pagesKobanana2 PDFSyifa Aulia NasutionNo ratings yet
- J. Agric. Food Chem. 2012, 60, 7204 7210Document7 pagesJ. Agric. Food Chem. 2012, 60, 7204 7210Duong Pham QuangNo ratings yet
- Persentasi Farfis HepniDocument28 pagesPersentasi Farfis HepniEpha Lumban GaolNo ratings yet
- International Journal of Pharma and Bio Sciences Issn 0975-6299Document10 pagesInternational Journal of Pharma and Bio Sciences Issn 0975-6299Narendrakumar GopakumaranNo ratings yet
- 1 s2.0 S2095311916615344 MainDocument7 pages1 s2.0 S2095311916615344 MainRosyida AdiniaNo ratings yet
- Alamar Blue Paper 2000Document6 pagesAlamar Blue Paper 2000deepak lourembamNo ratings yet
- JCPR 2016 8 5 879 888Document10 pagesJCPR 2016 8 5 879 888Lucía Ortega CabelloNo ratings yet
- The Bioactivity Study of Active Compounds in Bioactive SubstancesDocument9 pagesThe Bioactivity Study of Active Compounds in Bioactive SubstancesDinda Dwi SeptianiNo ratings yet
- Organochlorine Pesticides Residues in Mussels of Greek Island EviaDocument8 pagesOrganochlorine Pesticides Residues in Mussels of Greek Island Eviaandi tiaNo ratings yet
- Heydari 2015Document7 pagesHeydari 2015Phú Nguyễn VĩnhNo ratings yet
- Bacillus PolyfermenticusDocument6 pagesBacillus Polyfermenticussi275No ratings yet
- Cellulase Enzyme Production by Streptomyces SP Using Fruit Waste As SubstrateDocument5 pagesCellulase Enzyme Production by Streptomyces SP Using Fruit Waste As SubstrateHamka NurkayaNo ratings yet
- Biotechnology Journal - 2007 - Melissis - One Step Purification of Taq DNA Polymerase Using Nucleotide Mimetic AffinityDocument12 pagesBiotechnology Journal - 2007 - Melissis - One Step Purification of Taq DNA Polymerase Using Nucleotide Mimetic Affinity437guguNo ratings yet
- Analysis of Antioxidative Phenolic Compounds in ArtichokeDocument8 pagesAnalysis of Antioxidative Phenolic Compounds in ArtichokeAhmet UluadNo ratings yet
- Determination of Lawsone in Henna Powders by High Performance Thin Layer ChromatographyDocument6 pagesDetermination of Lawsone in Henna Powders by High Performance Thin Layer ChromatographynanoNo ratings yet
- DNAmetodo Aljanabi PDFDocument2 pagesDNAmetodo Aljanabi PDFCarlos Felix EscobedoNo ratings yet
- 13 - 15v5i2 - 4 Serratia Marcescens OU50TDocument5 pages13 - 15v5i2 - 4 Serratia Marcescens OU50TIsworo RukmiNo ratings yet
- Proteomic Analysis of The Venom of Heterometrus: Longimanus (Asian Black Scorpion)Document16 pagesProteomic Analysis of The Venom of Heterometrus: Longimanus (Asian Black Scorpion)Victor Isaac Perez soteloNo ratings yet
- Fraction A L EtOAc Soumise A La Diaion HP 20 - BDocument8 pagesFraction A L EtOAc Soumise A La Diaion HP 20 - BtangaNo ratings yet
- A Novel Multiparameter in Vitro ModelDocument9 pagesA Novel Multiparameter in Vitro ModelthyNo ratings yet
- Processes 08 00648 v2Document12 pagesProcesses 08 00648 v2Lidiane LimaNo ratings yet
- The Eco-Friendly Marine Gastropod Turbo Brunneus (L. 1758) and Its Vital Role in Future Pharmaceutical Industry Through GC-MS StudyDocument9 pagesThe Eco-Friendly Marine Gastropod Turbo Brunneus (L. 1758) and Its Vital Role in Future Pharmaceutical Industry Through GC-MS StudyEditor IJRITCCNo ratings yet
- Hsu2013 Ic50 Kuning JinggaDocument7 pagesHsu2013 Ic50 Kuning JinggaNuriNo ratings yet
- Purification and Properties of A Keratinolytic Metalloprotease From Microbacterium SPDocument10 pagesPurification and Properties of A Keratinolytic Metalloprotease From Microbacterium SPFaisal AwanNo ratings yet
- 39.03.16 International Journal of Biological Macromolecules 87 405-414 PDFDocument10 pages39.03.16 International Journal of Biological Macromolecules 87 405-414 PDFNarendrakumar GopakumaranNo ratings yet
- Box-Behnken Design Formulation Optimization and in Vitro Characterization of Rutin - Loaded Pla Nanoparticles Using Design-Expert SoftwareDocument13 pagesBox-Behnken Design Formulation Optimization and in Vitro Characterization of Rutin - Loaded Pla Nanoparticles Using Design-Expert SoftwareRAPPORTS DE PHARMACIENo ratings yet
- ADocument11 pagesARichie JuNo ratings yet
- Jurnal 13 PDFDocument5 pagesJurnal 13 PDFAri OktaviaNo ratings yet
- 150ChinJAnalChem2006 346 783-786Document7 pages150ChinJAnalChem2006 346 783-786milaNo ratings yet
- 2327 9834 Article p1449Document2 pages2327 9834 Article p1449Mpaz CastilloNo ratings yet
- Role of Plasmid in Production of Acetobacter Xylinum BiofilmsDocument4 pagesRole of Plasmid in Production of Acetobacter Xylinum BiofilmsShofiaNo ratings yet
- Journal of General MicrobiologyDocument14 pagesJournal of General MicrobiologykadriabdullahNo ratings yet
- A Cost-Effective RNA Extraction Technique From Animal Cells and Tissue Using Silica ColumnsDocument5 pagesA Cost-Effective RNA Extraction Technique From Animal Cells and Tissue Using Silica ColumnsNatalia Delgado VegaNo ratings yet
- DNA Comet Assay: A Simple Screening Technique For Identification of Some Irradiated FoodsDocument6 pagesDNA Comet Assay: A Simple Screening Technique For Identification of Some Irradiated FoodsIsael Maceo DíazNo ratings yet
- Elektroforesis KapilerDocument4 pagesElektroforesis KapilerRachma DyraNo ratings yet
- 30 2141 PDFDocument5 pages30 2141 PDFReanaldy Ibrahim Masudi PutraNo ratings yet
- 16 7 2019 IsolationDocument8 pages16 7 2019 IsolationNgan HoangNo ratings yet
- Effects of Micronization On The Physico-Chemical Properties of Peels of Three Root and Tuber CropsDocument5 pagesEffects of Micronization On The Physico-Chemical Properties of Peels of Three Root and Tuber CropsDadyan Raquel Flaquer FlorezNo ratings yet
- Phellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesDocument5 pagesPhellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesMinh NguyễnNo ratings yet
- Structural Features and Immunological Activity of A Polysaccharide From Dioscorea Opposita Thunb RootsDocument7 pagesStructural Features and Immunological Activity of A Polysaccharide From Dioscorea Opposita Thunb RootsFrontiersNo ratings yet
- Synthesis, Characterization, Cytotoxicity, DNA Cleavage and Antimicrobial Activity of Homodinuclear Lanthanide Complexes of Phenylthioacetic AcidDocument8 pagesSynthesis, Characterization, Cytotoxicity, DNA Cleavage and Antimicrobial Activity of Homodinuclear Lanthanide Complexes of Phenylthioacetic AcidABBS FEN REJI T. F.No ratings yet
- Cryoconcentration of Flavonoid Extract For Enhanced Biophotovoltaics and PH Sensitive Thin FilmsDocument12 pagesCryoconcentration of Flavonoid Extract For Enhanced Biophotovoltaics and PH Sensitive Thin Filmslina fernandaNo ratings yet
- Deteksi Molekuler 2Document10 pagesDeteksi Molekuler 2CHAYRA STUDIONo ratings yet
- Experimental Study of Electrostatic Precipitator Performance and Comparison With Existing Theoretical Prediction ModelsDocument23 pagesExperimental Study of Electrostatic Precipitator Performance and Comparison With Existing Theoretical Prediction ModelssamactrangNo ratings yet
- Lingerie Insight February 2011Document52 pagesLingerie Insight February 2011gab20100% (2)
- MLOG GX CMXA75 v4 0 322985a0 UM-ENDocument311 pagesMLOG GX CMXA75 v4 0 322985a0 UM-ENjamiekuangNo ratings yet
- Frizzell, 1987. Stress Orientation Determined From Fault Slip Data in Hampel Wash Area NevadaDocument10 pagesFrizzell, 1987. Stress Orientation Determined From Fault Slip Data in Hampel Wash Area Nevada黃詠晴No ratings yet
- Effects of Composition and Processing SEBS OIL PPDocument25 pagesEffects of Composition and Processing SEBS OIL PPBahadır Uğur AltunNo ratings yet
- QuadEquations PPT Alg2Document16 pagesQuadEquations PPT Alg2Kenny Ann Grace BatiancilaNo ratings yet
- Group Medical Insurance - 2020-21Document2 pagesGroup Medical Insurance - 2020-21Vilaz VijiNo ratings yet
- AREVA in NigerDocument20 pagesAREVA in Nigermushava nyokaNo ratings yet
- Sporlan TXV CatalogDocument40 pagesSporlan TXV CatalogDavid EllisNo ratings yet
- Pugh Method Example-2Document4 pagesPugh Method Example-2Shaikh AkhlaqueNo ratings yet
- The Cattell-Horn-Carroll Theory of Cognitive AbilitiesDocument13 pagesThe Cattell-Horn-Carroll Theory of Cognitive Abilitiessergio_poblete_ortegaNo ratings yet
- Manuel Resume 17Document2 pagesManuel Resume 17api-286125655No ratings yet
- UNIT-I (Sem-I) EVS - Environment & EcosystemDocument16 pagesUNIT-I (Sem-I) EVS - Environment & Ecosystemwalid ben aliNo ratings yet
- PDF Created With Pdffactory Pro Trial VersionDocument152 pagesPDF Created With Pdffactory Pro Trial Versionسري موتيك يوليانيNo ratings yet
- Case StudyDocument2 pagesCase StudyJoshua LaxamanaNo ratings yet
- Streamline Modular Electronic Sounder: Model SLM700Document2 pagesStreamline Modular Electronic Sounder: Model SLM700GersonDarioOrtizReyesNo ratings yet
- Research Compilation FinalDocument23 pagesResearch Compilation FinalJessa Mae LoredoNo ratings yet
- Surbir Singh Negi: Surbir99 - Negi@yahoo - Co.in, Surbir - Negi1@yahoo - Co.inDocument3 pagesSurbir Singh Negi: Surbir99 - Negi@yahoo - Co.in, Surbir - Negi1@yahoo - Co.insurbir_negi1100% (2)
- Biome Project RubricDocument1 pageBiome Project RubricshellyNo ratings yet
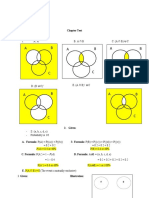
- A B A B A B: Chapter TestDocument2 pagesA B A B A B: Chapter TestgenieNo ratings yet
- The Importance of Mineral Elements For Humans, Domestic Animals and Plants: A ReviewDocument3 pagesThe Importance of Mineral Elements For Humans, Domestic Animals and Plants: A ReviewMaricel Canlas MacatoNo ratings yet
- Sika Backing FillerDocument2 pagesSika Backing FillerErdenebileg SangidorjNo ratings yet
- 2015 Packaging PresentationDocument56 pages2015 Packaging PresentationNayeli Mendez100% (1)
- SD Kelas 5Document2 pagesSD Kelas 5mts. miftahulhudaNo ratings yet
- The School As A Learning Environment.Document9 pagesThe School As A Learning Environment.Genevie Villahermosa QuirogaNo ratings yet
- Hero Rubric 2Document3 pagesHero Rubric 2api-293865917No ratings yet
- Machine Spindle Noses: 6 Bison - Bial S. ADocument2 pagesMachine Spindle Noses: 6 Bison - Bial S. AshanehatfieldNo ratings yet
- Historical Change and Ceramic Tradition: The Case of Macedonia - Zoi KotitsaDocument15 pagesHistorical Change and Ceramic Tradition: The Case of Macedonia - Zoi KotitsaSonjce Marceva100% (2)
- Employer BrandingDocument34 pagesEmployer BrandingTrinimafia100% (2)

























































































Download as pdf or txt
You might also like
- Balance Sheet & Income Statement For Hospital PDFDocument45 pagesBalance Sheet & Income Statement For Hospital PDFAdrianus PramudhitaNo ratings yet
- Kytococcus Sedentarius The Organism AssociatedDocument7 pagesKytococcus Sedentarius The Organism AssociatedMilena TomásNo ratings yet
- Antibacterial Principles From The Bark of Terminalia Arjuna: Scientific CorrespondenceDocument3 pagesAntibacterial Principles From The Bark of Terminalia Arjuna: Scientific CorrespondenceAshwani GaurNo ratings yet
- 1981 - Ekramoddoullah, Kisil, Sehon - Isolation of Allergenically Active Cytochrome C From Kentucky Blue Grass PollenDocument10 pages1981 - Ekramoddoullah, Kisil, Sehon - Isolation of Allergenically Active Cytochrome C From Kentucky Blue Grass Pollenpond_1993No ratings yet
- Nile Red - Kimura PDFDocument8 pagesNile Red - Kimura PDFSharif M Mizanur RahmanNo ratings yet
- Using Eclipta Prostrata (L.) Against Ht-29 Cell LineDocument5 pagesUsing Eclipta Prostrata (L.) Against Ht-29 Cell LineVardhana JanakiramanNo ratings yet
- Production, Purification and Characterization of An Extracellular Keratinase From Lysobacter NCIMB 9497Document6 pagesProduction, Purification and Characterization of An Extracellular Keratinase From Lysobacter NCIMB 9497Faisal AwanNo ratings yet
- EFRTDocument7 pagesEFRTDiego Fernando Jaimes GonzalezNo ratings yet
- Extraction and Characterization of Cellulose Nanocrystals From Corn StoverDocument7 pagesExtraction and Characterization of Cellulose Nanocrystals From Corn StoverBlessing HarvestNo ratings yet
- Structural Characterization of Cellulose ObtainedDocument5 pagesStructural Characterization of Cellulose ObtainedRezza MarcellaNo ratings yet
- Silva 2013Document4 pagesSilva 2013capau87No ratings yet
- Siedel 1984 Aquacultural-EngineeringDocument14 pagesSiedel 1984 Aquacultural-EngineeringJorge RodriguezNo ratings yet
- Binding and Uptake of Wheat Germ Agglutinin-Grafted PLGA-Nanospheres by Caco-2 MonolayersDocument7 pagesBinding and Uptake of Wheat Germ Agglutinin-Grafted PLGA-Nanospheres by Caco-2 MonolayerszeminkNo ratings yet
- Ravic Hand Ran 2016Document10 pagesRavic Hand Ran 2016Xuân LâmNo ratings yet
- WWW - Cjs 200902 0005Document8 pagesWWW - Cjs 200902 0005Uchiha AbdurrahmanNo ratings yet
- Pääbo Mitochondrial NAR 1988Document14 pagesPääbo Mitochondrial NAR 1988jaafari mohamedNo ratings yet
- Simultaneous Determination of Quinolones in Fish by Liquid Chromatography Coupled With Fluorescence Detection: Comparison of Sub-2 M Particles and Conventional C ColumnsDocument27 pagesSimultaneous Determination of Quinolones in Fish by Liquid Chromatography Coupled With Fluorescence Detection: Comparison of Sub-2 M Particles and Conventional C Columnsnguyen baoNo ratings yet
- Kobanana2 PDFDocument8 pagesKobanana2 PDFSyifa Aulia NasutionNo ratings yet
- J. Agric. Food Chem. 2012, 60, 7204 7210Document7 pagesJ. Agric. Food Chem. 2012, 60, 7204 7210Duong Pham QuangNo ratings yet
- Persentasi Farfis HepniDocument28 pagesPersentasi Farfis HepniEpha Lumban GaolNo ratings yet
- International Journal of Pharma and Bio Sciences Issn 0975-6299Document10 pagesInternational Journal of Pharma and Bio Sciences Issn 0975-6299Narendrakumar GopakumaranNo ratings yet
- 1 s2.0 S2095311916615344 MainDocument7 pages1 s2.0 S2095311916615344 MainRosyida AdiniaNo ratings yet
- Alamar Blue Paper 2000Document6 pagesAlamar Blue Paper 2000deepak lourembamNo ratings yet
- JCPR 2016 8 5 879 888Document10 pagesJCPR 2016 8 5 879 888Lucía Ortega CabelloNo ratings yet
- The Bioactivity Study of Active Compounds in Bioactive SubstancesDocument9 pagesThe Bioactivity Study of Active Compounds in Bioactive SubstancesDinda Dwi SeptianiNo ratings yet
- Organochlorine Pesticides Residues in Mussels of Greek Island EviaDocument8 pagesOrganochlorine Pesticides Residues in Mussels of Greek Island Eviaandi tiaNo ratings yet
- Heydari 2015Document7 pagesHeydari 2015Phú Nguyễn VĩnhNo ratings yet
- Bacillus PolyfermenticusDocument6 pagesBacillus Polyfermenticussi275No ratings yet
- Cellulase Enzyme Production by Streptomyces SP Using Fruit Waste As SubstrateDocument5 pagesCellulase Enzyme Production by Streptomyces SP Using Fruit Waste As SubstrateHamka NurkayaNo ratings yet
- Biotechnology Journal - 2007 - Melissis - One Step Purification of Taq DNA Polymerase Using Nucleotide Mimetic AffinityDocument12 pagesBiotechnology Journal - 2007 - Melissis - One Step Purification of Taq DNA Polymerase Using Nucleotide Mimetic Affinity437guguNo ratings yet
- Analysis of Antioxidative Phenolic Compounds in ArtichokeDocument8 pagesAnalysis of Antioxidative Phenolic Compounds in ArtichokeAhmet UluadNo ratings yet
- Determination of Lawsone in Henna Powders by High Performance Thin Layer ChromatographyDocument6 pagesDetermination of Lawsone in Henna Powders by High Performance Thin Layer ChromatographynanoNo ratings yet
- DNAmetodo Aljanabi PDFDocument2 pagesDNAmetodo Aljanabi PDFCarlos Felix EscobedoNo ratings yet
- 13 - 15v5i2 - 4 Serratia Marcescens OU50TDocument5 pages13 - 15v5i2 - 4 Serratia Marcescens OU50TIsworo RukmiNo ratings yet
- Proteomic Analysis of The Venom of Heterometrus: Longimanus (Asian Black Scorpion)Document16 pagesProteomic Analysis of The Venom of Heterometrus: Longimanus (Asian Black Scorpion)Victor Isaac Perez soteloNo ratings yet
- Fraction A L EtOAc Soumise A La Diaion HP 20 - BDocument8 pagesFraction A L EtOAc Soumise A La Diaion HP 20 - BtangaNo ratings yet
- A Novel Multiparameter in Vitro ModelDocument9 pagesA Novel Multiparameter in Vitro ModelthyNo ratings yet
- Processes 08 00648 v2Document12 pagesProcesses 08 00648 v2Lidiane LimaNo ratings yet
- The Eco-Friendly Marine Gastropod Turbo Brunneus (L. 1758) and Its Vital Role in Future Pharmaceutical Industry Through GC-MS StudyDocument9 pagesThe Eco-Friendly Marine Gastropod Turbo Brunneus (L. 1758) and Its Vital Role in Future Pharmaceutical Industry Through GC-MS StudyEditor IJRITCCNo ratings yet
- Hsu2013 Ic50 Kuning JinggaDocument7 pagesHsu2013 Ic50 Kuning JinggaNuriNo ratings yet
- Purification and Properties of A Keratinolytic Metalloprotease From Microbacterium SPDocument10 pagesPurification and Properties of A Keratinolytic Metalloprotease From Microbacterium SPFaisal AwanNo ratings yet
- 39.03.16 International Journal of Biological Macromolecules 87 405-414 PDFDocument10 pages39.03.16 International Journal of Biological Macromolecules 87 405-414 PDFNarendrakumar GopakumaranNo ratings yet
- Box-Behnken Design Formulation Optimization and in Vitro Characterization of Rutin - Loaded Pla Nanoparticles Using Design-Expert SoftwareDocument13 pagesBox-Behnken Design Formulation Optimization and in Vitro Characterization of Rutin - Loaded Pla Nanoparticles Using Design-Expert SoftwareRAPPORTS DE PHARMACIENo ratings yet
- ADocument11 pagesARichie JuNo ratings yet
- Jurnal 13 PDFDocument5 pagesJurnal 13 PDFAri OktaviaNo ratings yet
- 150ChinJAnalChem2006 346 783-786Document7 pages150ChinJAnalChem2006 346 783-786milaNo ratings yet
- 2327 9834 Article p1449Document2 pages2327 9834 Article p1449Mpaz CastilloNo ratings yet
- Role of Plasmid in Production of Acetobacter Xylinum BiofilmsDocument4 pagesRole of Plasmid in Production of Acetobacter Xylinum BiofilmsShofiaNo ratings yet
- Journal of General MicrobiologyDocument14 pagesJournal of General MicrobiologykadriabdullahNo ratings yet
- A Cost-Effective RNA Extraction Technique From Animal Cells and Tissue Using Silica ColumnsDocument5 pagesA Cost-Effective RNA Extraction Technique From Animal Cells and Tissue Using Silica ColumnsNatalia Delgado VegaNo ratings yet
- DNA Comet Assay: A Simple Screening Technique For Identification of Some Irradiated FoodsDocument6 pagesDNA Comet Assay: A Simple Screening Technique For Identification of Some Irradiated FoodsIsael Maceo DíazNo ratings yet
- Elektroforesis KapilerDocument4 pagesElektroforesis KapilerRachma DyraNo ratings yet
- 30 2141 PDFDocument5 pages30 2141 PDFReanaldy Ibrahim Masudi PutraNo ratings yet
- 16 7 2019 IsolationDocument8 pages16 7 2019 IsolationNgan HoangNo ratings yet
- Effects of Micronization On The Physico-Chemical Properties of Peels of Three Root and Tuber CropsDocument5 pagesEffects of Micronization On The Physico-Chemical Properties of Peels of Three Root and Tuber CropsDadyan Raquel Flaquer FlorezNo ratings yet
- Phellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesDocument5 pagesPhellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesMinh NguyễnNo ratings yet
- Structural Features and Immunological Activity of A Polysaccharide From Dioscorea Opposita Thunb RootsDocument7 pagesStructural Features and Immunological Activity of A Polysaccharide From Dioscorea Opposita Thunb RootsFrontiersNo ratings yet
- Synthesis, Characterization, Cytotoxicity, DNA Cleavage and Antimicrobial Activity of Homodinuclear Lanthanide Complexes of Phenylthioacetic AcidDocument8 pagesSynthesis, Characterization, Cytotoxicity, DNA Cleavage and Antimicrobial Activity of Homodinuclear Lanthanide Complexes of Phenylthioacetic AcidABBS FEN REJI T. F.No ratings yet
- Cryoconcentration of Flavonoid Extract For Enhanced Biophotovoltaics and PH Sensitive Thin FilmsDocument12 pagesCryoconcentration of Flavonoid Extract For Enhanced Biophotovoltaics and PH Sensitive Thin Filmslina fernandaNo ratings yet
- Deteksi Molekuler 2Document10 pagesDeteksi Molekuler 2CHAYRA STUDIONo ratings yet
- Experimental Study of Electrostatic Precipitator Performance and Comparison With Existing Theoretical Prediction ModelsDocument23 pagesExperimental Study of Electrostatic Precipitator Performance and Comparison With Existing Theoretical Prediction ModelssamactrangNo ratings yet
- Lingerie Insight February 2011Document52 pagesLingerie Insight February 2011gab20100% (2)
- MLOG GX CMXA75 v4 0 322985a0 UM-ENDocument311 pagesMLOG GX CMXA75 v4 0 322985a0 UM-ENjamiekuangNo ratings yet
- Frizzell, 1987. Stress Orientation Determined From Fault Slip Data in Hampel Wash Area NevadaDocument10 pagesFrizzell, 1987. Stress Orientation Determined From Fault Slip Data in Hampel Wash Area Nevada黃詠晴No ratings yet
- Effects of Composition and Processing SEBS OIL PPDocument25 pagesEffects of Composition and Processing SEBS OIL PPBahadır Uğur AltunNo ratings yet
- QuadEquations PPT Alg2Document16 pagesQuadEquations PPT Alg2Kenny Ann Grace BatiancilaNo ratings yet
- Group Medical Insurance - 2020-21Document2 pagesGroup Medical Insurance - 2020-21Vilaz VijiNo ratings yet
- AREVA in NigerDocument20 pagesAREVA in Nigermushava nyokaNo ratings yet
- Sporlan TXV CatalogDocument40 pagesSporlan TXV CatalogDavid EllisNo ratings yet
- Pugh Method Example-2Document4 pagesPugh Method Example-2Shaikh AkhlaqueNo ratings yet
- The Cattell-Horn-Carroll Theory of Cognitive AbilitiesDocument13 pagesThe Cattell-Horn-Carroll Theory of Cognitive Abilitiessergio_poblete_ortegaNo ratings yet
- Manuel Resume 17Document2 pagesManuel Resume 17api-286125655No ratings yet
- UNIT-I (Sem-I) EVS - Environment & EcosystemDocument16 pagesUNIT-I (Sem-I) EVS - Environment & Ecosystemwalid ben aliNo ratings yet
- PDF Created With Pdffactory Pro Trial VersionDocument152 pagesPDF Created With Pdffactory Pro Trial Versionسري موتيك يوليانيNo ratings yet
- Case StudyDocument2 pagesCase StudyJoshua LaxamanaNo ratings yet
- Streamline Modular Electronic Sounder: Model SLM700Document2 pagesStreamline Modular Electronic Sounder: Model SLM700GersonDarioOrtizReyesNo ratings yet
- Research Compilation FinalDocument23 pagesResearch Compilation FinalJessa Mae LoredoNo ratings yet
- Surbir Singh Negi: Surbir99 - Negi@yahoo - Co.in, Surbir - Negi1@yahoo - Co.inDocument3 pagesSurbir Singh Negi: Surbir99 - Negi@yahoo - Co.in, Surbir - Negi1@yahoo - Co.insurbir_negi1100% (2)
- Biome Project RubricDocument1 pageBiome Project RubricshellyNo ratings yet
- A B A B A B: Chapter TestDocument2 pagesA B A B A B: Chapter TestgenieNo ratings yet
- The Importance of Mineral Elements For Humans, Domestic Animals and Plants: A ReviewDocument3 pagesThe Importance of Mineral Elements For Humans, Domestic Animals and Plants: A ReviewMaricel Canlas MacatoNo ratings yet
- Sika Backing FillerDocument2 pagesSika Backing FillerErdenebileg SangidorjNo ratings yet
- 2015 Packaging PresentationDocument56 pages2015 Packaging PresentationNayeli Mendez100% (1)
- SD Kelas 5Document2 pagesSD Kelas 5mts. miftahulhudaNo ratings yet
- The School As A Learning Environment.Document9 pagesThe School As A Learning Environment.Genevie Villahermosa QuirogaNo ratings yet
- Hero Rubric 2Document3 pagesHero Rubric 2api-293865917No ratings yet
- Machine Spindle Noses: 6 Bison - Bial S. ADocument2 pagesMachine Spindle Noses: 6 Bison - Bial S. AshanehatfieldNo ratings yet
- Historical Change and Ceramic Tradition: The Case of Macedonia - Zoi KotitsaDocument15 pagesHistorical Change and Ceramic Tradition: The Case of Macedonia - Zoi KotitsaSonjce Marceva100% (2)
- Employer BrandingDocument34 pagesEmployer BrandingTrinimafia100% (2)