Download as pdf or txt
You might also like
- BCMB 415 Exam 3, Fall 2016 KeyDocument9 pagesBCMB 415 Exam 3, Fall 2016 KeyzzmasterNo ratings yet
- Arranging Fine Perfume CompositionsDocument338 pagesArranging Fine Perfume CompositionsAndré Mattos100% (1)
- The Extraordinary Channels (PDFDrive)Document28 pagesThe Extraordinary Channels (PDFDrive)Christos Golobias100% (4)
- Review Literature: Sien Lin. Et.,al in His Study He Had Developed Chitosan Scaffolds CombinedDocument4 pagesReview Literature: Sien Lin. Et.,al in His Study He Had Developed Chitosan Scaffolds Combinedsree anugraphicsNo ratings yet
- Food Chemistry: Sittichoke Sinthusamran, Soottawat Benjakul, Hideki KishimuraDocument7 pagesFood Chemistry: Sittichoke Sinthusamran, Soottawat Benjakul, Hideki KishimuraAmi UtamiNo ratings yet
- Isolation and Characterization of Thermostable Collagen From The Marine Eel-Fish (Evenchelys Macrura)Document11 pagesIsolation and Characterization of Thermostable Collagen From The Marine Eel-Fish (Evenchelys Macrura)Hartanty EntedingNo ratings yet
- Efeitos Da Ingestão de Peptídeos de ColágenoDocument5 pagesEfeitos Da Ingestão de Peptídeos de ColágenoAdriana VasconcelosNo ratings yet
- Collagen Extraction Process4Document8 pagesCollagen Extraction Process4gandurikNo ratings yet
- Ovine Tendon Collagen Extraction, Characterization and Fabrication of Thin Films For Tissue EngineeringDocument9 pagesOvine Tendon Collagen Extraction, Characterization and Fabrication of Thin Films For Tissue EngineeringGuilherme de OliveiraNo ratings yet
- Raman 2014Document8 pagesRaman 2014iulianaNo ratings yet
- Tsiplakou Et Al., 2018-LDocument11 pagesTsiplakou Et Al., 2018-LLuis Alberto Canul KuNo ratings yet
- Food Bioscience: Qiang Zhang, Qianqian Wang, Shun LV, Jianfeng Lu, Shaotong Jiang, Joe M. Regenstein, Lin LinDocument8 pagesFood Bioscience: Qiang Zhang, Qianqian Wang, Shun LV, Jianfeng Lu, Shaotong Jiang, Joe M. Regenstein, Lin LinsillysiswaniNo ratings yet
- Comparative Study Between Different MethodsDocument4 pagesComparative Study Between Different MethodsNajat AyubNo ratings yet
- Isolation and Characterization of Collagen From Fish Waste Material-Skin, Scales and Fins of Catla Catla and Cirrhinus MrigalaDocument10 pagesIsolation and Characterization of Collagen From Fish Waste Material-Skin, Scales and Fins of Catla Catla and Cirrhinus MrigalaIrpan RomadonaNo ratings yet
- BacilluscereuscollagenaseDocument10 pagesBacilluscereuscollagenasealiNo ratings yet
- Collagen Extraction ProcessDocument18 pagesCollagen Extraction ProcessDhita PramestyaNo ratings yet
- Application of Marine Collagen For Stem Cell Based - Chao LiuDocument10 pagesApplication of Marine Collagen For Stem Cell Based - Chao Liukosmetika GMSNo ratings yet
- Tan y Chang 2018Document10 pagesTan y Chang 2018David SalirrosasNo ratings yet
- Assessment of Season-Dependent Body Condition Scores in Relation To Faecal Glucocorticoid Metabolites in Free-Ranging Asian ElephantsDocument14 pagesAssessment of Season-Dependent Body Condition Scores in Relation To Faecal Glucocorticoid Metabolites in Free-Ranging Asian ElephantsDede Aulia RahmanNo ratings yet
- Viability Test of Fish Scales Collagen From Oshphronemus Gouramy On Osteoblast Cell CultureDocument5 pagesViability Test of Fish Scales Collagen From Oshphronemus Gouramy On Osteoblast Cell CultureRAKERNAS PDGI XINo ratings yet
- Sip Additional InfoDocument9 pagesSip Additional Infogracelyn a. alejoNo ratings yet
- Chen 2019Document9 pagesChen 2019Nur DonaNo ratings yet
- High-Throughput Physically Based Approach For Mammalian Cell EncapsulationDocument6 pagesHigh-Throughput Physically Based Approach For Mammalian Cell Encapsulation健康新體驗No ratings yet
- Ecnu 13 00529Document16 pagesEcnu 13 00529Svetlana Khyn SeseNo ratings yet
- Microfluidic Gut-On-A-Chip With Three-Dimensional Villi StructureDocument10 pagesMicrofluidic Gut-On-A-Chip With Three-Dimensional Villi StructureThamiris SouzaNo ratings yet
- (Colageno) Chattopadhyay - Et - Al-2014-Biopolymers PDFDocument13 pages(Colageno) Chattopadhyay - Et - Al-2014-Biopolymers PDFAndré Zamith SelvaggioNo ratings yet
- Collagen From Sea Cucumber (Stichopus Variegatus) As An Alternative Source of Halal CollagenDocument3 pagesCollagen From Sea Cucumber (Stichopus Variegatus) As An Alternative Source of Halal CollagenDony HermantoNo ratings yet
- Collagen in Food and Beverage Industries Minireview: International Food Research Journal 22 (1) : 1 - 8 (2015)Document8 pagesCollagen in Food and Beverage Industries Minireview: International Food Research Journal 22 (1) : 1 - 8 (2015)Dhiqbull GembulNo ratings yet
- 10.1007/s00441 014 1938 1Document9 pages10.1007/s00441 014 1938 1Widi Marsha FadilaNo ratings yet
- Isolation Characterization and Molecular Weight Determination of Collagen From Marine Sponge Spirastrella Inconstans (Dendy)Document8 pagesIsolation Characterization and Molecular Weight Determination of Collagen From Marine Sponge Spirastrella Inconstans (Dendy)Paulo JuniorNo ratings yet
- Tugas 6Document5 pagesTugas 6faris naufalNo ratings yet
- 7.biochemical and Structural Characterization of Sturgeon FishDocument10 pages7.biochemical and Structural Characterization of Sturgeon FishJonathanCoralNo ratings yet
- IFRO v19n2p768 enDocument12 pagesIFRO v19n2p768 enHarold PanganoronNo ratings yet
- Aloevera PDFDocument9 pagesAloevera PDFDeimer OrtizNo ratings yet
- Final VersionDocument11 pagesFinal Versionariw39596No ratings yet
- Chitosan-Alginate Microcapsules For Oral Delivery of Egg YolkDocument8 pagesChitosan-Alginate Microcapsules For Oral Delivery of Egg YolkRo BellingeriNo ratings yet
- Sustainable Chemistry and PharmacyDocument8 pagesSustainable Chemistry and PharmacyDavid SalirrosasNo ratings yet
- Fabrication of Human Hair Keratin-Jellyfish Collagen-Eggshell-Derived HA For Bone Tissue RegDocument34 pagesFabrication of Human Hair Keratin-Jellyfish Collagen-Eggshell-Derived HA For Bone Tissue RegAlina ComanNo ratings yet
- Bioresource Technology: Ankita Juneja, Frank W.R. Chaplen, Ganti S. MurthyDocument8 pagesBioresource Technology: Ankita Juneja, Frank W.R. Chaplen, Ganti S. MurthyMermaidNo ratings yet
- Characterization of Gelatin From The Skin of Farmed Amur Sturgeon Acipenser SchrenckiiDocument11 pagesCharacterization of Gelatin From The Skin of Farmed Amur Sturgeon Acipenser SchrenckiiDanielNo ratings yet
- Taxonomic and Functional Assessment Using Metatranscriptomics Reveals The Effect of Angus Cattle On Rumen Microbial SignaturesDocument15 pagesTaxonomic and Functional Assessment Using Metatranscriptomics Reveals The Effect of Angus Cattle On Rumen Microbial SignaturesAndré Luis NevesNo ratings yet
- Histological Structure and Chemical Composition of Farmed Giant Catfish's SkinDocument5 pagesHistological Structure and Chemical Composition of Farmed Giant Catfish's SkinVo Dong Phuong100% (1)
- Genome-Scale MetabolicDocument6 pagesGenome-Scale MetabolicMohamed BahgatNo ratings yet
- Biophysicochemical Evaluation of ChitosaDocument10 pagesBiophysicochemical Evaluation of ChitosaGraciele MariaNo ratings yet
- Collagen Extraction Process2Document10 pagesCollagen Extraction Process2gandurikNo ratings yet
- IJMLR221701Document11 pagesIJMLR221701sandeep raiNo ratings yet
- 3-D Printed Meat Alternatives Based On Pea and Single Cell Proteins and 3Document11 pages3-D Printed Meat Alternatives Based On Pea and Single Cell Proteins and 3079 Niha.sNo ratings yet
- Collagen Hydrolysates Modulate Cartilage MetabolismDocument14 pagesCollagen Hydrolysates Modulate Cartilage MetabolismНадія АсадуллаєваNo ratings yet
- Hydrolyzed Collagen-Sources and ApplicationsDocument16 pagesHydrolyzed Collagen-Sources and ApplicationsImelda Irma IsauraNo ratings yet
- Polymers: in Vitro Degradation of Electrospun Poly (Lactic-Co-Glycolic Acid) (PLGA) For Oral Mucosa RegenerationDocument19 pagesPolymers: in Vitro Degradation of Electrospun Poly (Lactic-Co-Glycolic Acid) (PLGA) For Oral Mucosa RegenerationDiego Caproni de MoraisNo ratings yet
- Characterization and Controlled Release Aloe Extract of Collagen Protein Modified Cotton FiberDocument7 pagesCharacterization and Controlled Release Aloe Extract of Collagen Protein Modified Cotton FiberVanessaCamposNo ratings yet
- Sharmila 2020Document8 pagesSharmila 2020chemistrycsir5No ratings yet
- In Vitro Engineering of Human Autogenous Cartilage: Ursula Anderer and Jeanette LiberaDocument10 pagesIn Vitro Engineering of Human Autogenous Cartilage: Ursula Anderer and Jeanette LiberaANIS MURNIATINo ratings yet
- Afifah 2019 IOP Conf. Ser. Earth Environ. Sci. 335 012031Document10 pagesAfifah 2019 IOP Conf. Ser. Earth Environ. Sci. 335 012031nurul auliaNo ratings yet
- Collagen ExtarctionDocument5 pagesCollagen ExtarctionAlmiraNadiaNo ratings yet
- Comparison of The Structural Characteristics of Native Collagen Fibrils Derived From Bovine Tendons Using Two Different Methods, Modified Acid Solubilized and Pepsin Aided ExtractionDocument13 pagesComparison of The Structural Characteristics of Native Collagen Fibrils Derived From Bovine Tendons Using Two Different Methods, Modified Acid Solubilized and Pepsin Aided Extractiononur ateşNo ratings yet
- Los Polifenoles Del Cacao Se Absorben en El Modelo Celular de Epitelio Intestinal Caco-2Document7 pagesLos Polifenoles Del Cacao Se Absorben en El Modelo Celular de Epitelio Intestinal Caco-2BrayanTrujilloNo ratings yet
- Short Communication: Dietary Bovine Milk-Derived Exosomes: Improve Bone Health in An Osteoporosis-Induced Mouse ModelDocument9 pagesShort Communication: Dietary Bovine Milk-Derived Exosomes: Improve Bone Health in An Osteoporosis-Induced Mouse ModelIneke NabilahNo ratings yet
- Impact of Source and Manufacturing of CollagenDocument14 pagesImpact of Source and Manufacturing of CollagenjcastNo ratings yet
- Bovine Osteoblasts Cultured On Polyanionic Collagen Scaffolds: An Ultrastructural and Immunocytochemical StudyDocument10 pagesBovine Osteoblasts Cultured On Polyanionic Collagen Scaffolds: An Ultrastructural and Immunocytochemical StudystaseekNo ratings yet
- Biosintesa GlukomananDocument12 pagesBiosintesa GlukomananDetya Pitaloka SariNo ratings yet
- J Cellular Molecular Medi - 2013 - Maurel - Effect of The Alcohol Consumption On Osteocyte Cell Processes A MolecularDocument14 pagesJ Cellular Molecular Medi - 2013 - Maurel - Effect of The Alcohol Consumption On Osteocyte Cell Processes A MolecularSaffa AzharaaniNo ratings yet
- Success Factors for Fish Larval ProductionFrom EverandSuccess Factors for Fish Larval ProductionLuis ConceicaoNo ratings yet
- Ada DH LumayanDocument16 pagesAda DH LumayanwindaNo ratings yet
- Hema Efek Dari HidrolisisDocument9 pagesHema Efek Dari HidrolisiswindaNo ratings yet
- ABG10Document5 pagesABG10windaNo ratings yet
- The Influence of Solvent Concentration and Long Soaking To Quality Gelatin Fish Scales Red Snapper (Lutjanus Camphecanus SP.)Document6 pagesThe Influence of Solvent Concentration and Long Soaking To Quality Gelatin Fish Scales Red Snapper (Lutjanus Camphecanus SP.)windaNo ratings yet
- LWT - Food Science and TechnologyDocument8 pagesLWT - Food Science and TechnologywindaNo ratings yet
- Extraction of Fish Oil From Parexocoetus Brachypterus (Flying Fish) Via Soxhlet Extraction MethodDocument7 pagesExtraction of Fish Oil From Parexocoetus Brachypterus (Flying Fish) Via Soxhlet Extraction MethodwindaNo ratings yet
- Ferdos TongkolDocument11 pagesFerdos TongkolwindaNo ratings yet
- Development of A Nutritionally Enriched Fish Cake From Mixed Fish SpeciesDocument7 pagesDevelopment of A Nutritionally Enriched Fish Cake From Mixed Fish SpecieswindaNo ratings yet
- Fatty-Acid Water : Content of OceanDocument10 pagesFatty-Acid Water : Content of OceanwindaNo ratings yet
- Sid Meyer's Alpha Centauri Strategy GuideDocument112 pagesSid Meyer's Alpha Centauri Strategy GuideOndřej PovolnýNo ratings yet
- To Compare Rate of FermentationDocument12 pagesTo Compare Rate of FermentationSahilKumar33% (3)
- Class 8 Nso PaperDocument9 pagesClass 8 Nso PaperYatish Goyal75% (4)
- Targeted Muscle Reinnervation A Paradigm Shift.3Document9 pagesTargeted Muscle Reinnervation A Paradigm Shift.3TRAUMATOLOGIA HEGNo ratings yet
- 01B-Arah Dan Bidang KristalDocument20 pages01B-Arah Dan Bidang KristalDyah Ayu DaratikaNo ratings yet
- Answer Sheet Phil IriDocument4 pagesAnswer Sheet Phil IriEiron Almeron100% (2)
- Application of Nanotechnology in Medical FieldDocument9 pagesApplication of Nanotechnology in Medical FieldKyleigh HaschkeNo ratings yet
- Healing TraumaDocument12 pagesHealing Traumaana.paula.a.martins100% (1)
- Culturing Beneficial Microbiology For Farming PDFDocument28 pagesCulturing Beneficial Microbiology For Farming PDFntv2000100% (1)
- Science 5 Sample Papers With SolutionsDocument52 pagesScience 5 Sample Papers With SolutionsAryaman TiwariNo ratings yet
- Child and Adolescent Learner and Learning Principles: College of EducationDocument36 pagesChild and Adolescent Learner and Learning Principles: College of EducationDestiny Jon Ivan100% (2)
- Linking Family Hardship To Children's LivesDocument16 pagesLinking Family Hardship To Children's LivesGabrielLopezNo ratings yet
- iBT TOEFL 托福词汇 词以类记Document15 pagesiBT TOEFL 托福词汇 词以类记beat2013No ratings yet
- Expression of Biological InformationDocument7 pagesExpression of Biological InformationNur Amalina KhozariNo ratings yet
- 考前阅读真题3Document17 pages考前阅读真题3gaoyuying73No ratings yet
- Journal Pre-Proofs: Food ChemistryDocument29 pagesJournal Pre-Proofs: Food ChemistrySubaraDeniNo ratings yet
- Animal Classification Phylogeny and Organization 1Document17 pagesAnimal Classification Phylogeny and Organization 1api-28507886575% (4)
- How Successful People Handle Toxic People - ForbesDocument6 pagesHow Successful People Handle Toxic People - ForbesRick HunterNo ratings yet
- HBSF Unit 1 Slides (Set 2)Document19 pagesHBSF Unit 1 Slides (Set 2)Jesus Juan ChavoyaNo ratings yet
- Anti-Plaque and Anti-Gingivitis Effect of Papain, Bromelain, Miswak and Neem Containing Dentifrice: A Randomized Controlled TrialDocument5 pagesAnti-Plaque and Anti-Gingivitis Effect of Papain, Bromelain, Miswak and Neem Containing Dentifrice: A Randomized Controlled TrialKIH 20162017No ratings yet
- Module 1 - Introduction To EcologyDocument6 pagesModule 1 - Introduction To EcologyNelzen GarayNo ratings yet
- Quality Control Charts in Hematology: HemoglobinDocument2 pagesQuality Control Charts in Hematology: HemoglobinDayledaniel SorvetoNo ratings yet
- Maternal & Child Health Nursing Flashcards QuizletDocument1 pageMaternal & Child Health Nursing Flashcards QuizletJosh PagnamitanNo ratings yet
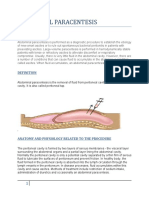
- Abdominal ParacentesisDocument14 pagesAbdominal ParacentesisManisha Thakur100% (1)
- V Alagarsamy: As Per The Latest Syllabus Prescribed by Pharmacy Council of IndiaDocument13 pagesV Alagarsamy: As Per The Latest Syllabus Prescribed by Pharmacy Council of IndiaPriyanshiNo ratings yet
- Antidepressive Effects of Ginsenoside Rg1 Via Regulation of HPA and HPG AxisDocument10 pagesAntidepressive Effects of Ginsenoside Rg1 Via Regulation of HPA and HPG AxisMylena SilvaNo ratings yet
- Pituitary and Hypothalamus: The Body's Control Center: Lesson A2.2Document40 pagesPituitary and Hypothalamus: The Body's Control Center: Lesson A2.2Khobie PabilicoNo ratings yet