
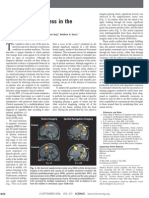
Referencia 39 2021 Kitazawa Mutacion Adrb1-A187v
Referencia 39 2021 Kitazawa Mutacion Adrb1-A187v
You might also like
- F& C OSCE Notes - University of Manchester, WL GanDocument57 pagesF& C OSCE Notes - University of Manchester, WL GanWeh Loong Gan100% (7)
- Neurofibromin 1 Regulates Early Developmental Sleep in DrosophilaDocument9 pagesNeurofibromin 1 Regulates Early Developmental Sleep in DrosophilaFernandoNo ratings yet
- A. Basic Sleep Science V. Arousal and BehaviorDocument2 pagesA. Basic Sleep Science V. Arousal and BehaviorivanNo ratings yet
- SleepDocument9 pagesSleepLobnaNo ratings yet
- Benefits of Napping and An Extended Duration of Recovery Sleep On Alertness and Immune Cells After Acute Sleep RestrictionDocument9 pagesBenefits of Napping and An Extended Duration of Recovery Sleep On Alertness and Immune Cells After Acute Sleep RestrictionMatheus VilarinhoNo ratings yet
- 1 s2.0 S0896627313006533 MainDocument13 pages1 s2.0 S0896627313006533 Mainshriram1082883No ratings yet
- Cho 2013Document4 pagesCho 2013Kassandra Alfaro AlvarezNo ratings yet
- Sleepdisturbanceslinked Togeneticdisorders: Rimawati TedjasukmanaDocument10 pagesSleepdisturbanceslinked Togeneticdisorders: Rimawati TedjasukmanaLeilybadryaNo ratings yet
- Immanuel 2014Document4 pagesImmanuel 2014Mary Alarcon PalominoNo ratings yet
- Sleep Paralysis: June 2012Document8 pagesSleep Paralysis: June 2012renuka1011No ratings yet
- SCIENCE 2006 Detecting Awareness in The Vegetative StateDocument1 pageSCIENCE 2006 Detecting Awareness in The Vegetative StatepreghieraperlavitaNo ratings yet
- Creatine Supplementation Reduces Sleep Need and Homeostatic Sleep Pressure in RatsDocument9 pagesCreatine Supplementation Reduces Sleep Need and Homeostatic Sleep Pressure in RatswNo ratings yet
- Parasonias em REMDocument16 pagesParasonias em REMhcrislopes5617No ratings yet
- Sleep Medicine: Original ArticleDocument11 pagesSleep Medicine: Original ArticleaviyahnikmahNo ratings yet
- Are There Parallel Channels in The Vestibular Nerve?: Ellengene H. PetersonDocument8 pagesAre There Parallel Channels in The Vestibular Nerve?: Ellengene H. PetersonramopavelNo ratings yet
- Sleep Paralysis: June 2012Document8 pagesSleep Paralysis: June 2012edamwone04No ratings yet
- 1 s2.0 S1095643321002221 Main 1Document12 pages1 s2.0 S1095643321002221 Main 11603365No ratings yet
- Human Neurobiolog y Sleep Sleep DeprivatDocument6 pagesHuman Neurobiolog y Sleep Sleep Deprivatmelgarhoward90No ratings yet
- Neurostimulation-techniques-to-enhance-sleep-and-improve-cognition-in-agingDocument16 pagesNeurostimulation-techniques-to-enhance-sleep-and-improve-cognition-in-agingPrakash N KNo ratings yet
- 523 FullDocument3 pages523 FulldesckygamerNo ratings yet
- Sleep in Gerstmann-Straüssler-Scheinker Disease.Document5 pagesSleep in Gerstmann-Straüssler-Scheinker Disease.Lídia YoshiharaNo ratings yet
- Vision Research: David H. Mcdougal, Paul D. GamlinDocument16 pagesVision Research: David H. Mcdougal, Paul D. GamlinCristina GómezNo ratings yet
- Sleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerriDocument9 pagesSleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerripsicosmosNo ratings yet
- The Importance of Astrocyte-Derived Purines in The Modulation of SleepDocument11 pagesThe Importance of Astrocyte-Derived Purines in The Modulation of Sleepapi-214563130No ratings yet
- Borbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Document7 pagesBorbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Sebastian PereiraNo ratings yet
- Edwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitsDocument16 pagesEdwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitssbachorickNo ratings yet
- Short Sleep - Poor Sleep? A Polysomnographic Study in A Large Population Based Sample of WomenDocument7 pagesShort Sleep - Poor Sleep? A Polysomnographic Study in A Large Population Based Sample of Womenmahfuz emonNo ratings yet
- Homer1a Is A Core Brain Molecular Correlate of Sleep LossDocument6 pagesHomer1a Is A Core Brain Molecular Correlate of Sleep LosswestNo ratings yet
- Sleep and Wakefulness in Drosophila MelanogasterDocument7 pagesSleep and Wakefulness in Drosophila MelanogasterFernandoNo ratings yet
- Astrocito y SueñoDocument17 pagesAstrocito y SueñoLEISLY TATIANA OVIEDO GOMEZNo ratings yet
- Extended Wakefulness: Compromised Metabolics in and Degeneration of Locus Ceruleus NeuronsDocument14 pagesExtended Wakefulness: Compromised Metabolics in and Degeneration of Locus Ceruleus NeuronsRehaan FayazNo ratings yet
- Des Car GableDocument10 pagesDes Car GableDago CastilloNo ratings yet
- HHS Public Access: Impact of Sleep Disturbances On Neurodegeneration: Insight From Studies in Animal ModelsDocument25 pagesHHS Public Access: Impact of Sleep Disturbances On Neurodegeneration: Insight From Studies in Animal ModelsPawan MishraNo ratings yet
- A Role For Non-Rapid-Eye-Movement Sleep Homeostasis in Perceptual LearningDocument7 pagesA Role For Non-Rapid-Eye-Movement Sleep Homeostasis in Perceptual LearningPravin KumarNo ratings yet
- Sleep in Drosophila and Its ContextDocument19 pagesSleep in Drosophila and Its ContextFernandoNo ratings yet
- 2001 Atlas, Rules, and Recording Techniques For The Scoring of Cyclic Alternating Pattern (CAP) in Human SleepDocument17 pages2001 Atlas, Rules, and Recording Techniques For The Scoring of Cyclic Alternating Pattern (CAP) in Human SleepNHUNG Hoàng HuyềnNo ratings yet
- Epilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CookDocument11 pagesEpilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CooknurhikmahNo ratings yet
- Manipulations of Amyloid Precursor Protein Cleavage Disrupt The Circadian Clock in Aging DrosophilaDocument10 pagesManipulations of Amyloid Precursor Protein Cleavage Disrupt The Circadian Clock in Aging DrosophilaRan LinNo ratings yet
- Sleep 26 2 117Document10 pagesSleep 26 2 117ClaudiaNo ratings yet
- Nocturnal Light Exposure Impairs Affective Responses in A Wavelength-Dependent MannerDocument7 pagesNocturnal Light Exposure Impairs Affective Responses in A Wavelength-Dependent MannerLiana DimaNo ratings yet
- 1 s2.0 S0167876016304329 MainDocument1 page1 s2.0 S0167876016304329 Mainmincuandrada10No ratings yet
- Whitehead 2018 Emergence of Mature Cortical Activity in Wakefulness and Sleep in Healthy Preterm and Full Term InfantsDocument9 pagesWhitehead 2018 Emergence of Mature Cortical Activity in Wakefulness and Sleep in Healthy Preterm and Full Term InfantsC SNo ratings yet
- Baird NeuroImage 16 1120 2002 PDFDocument7 pagesBaird NeuroImage 16 1120 2002 PDFVezér RóbertNo ratings yet
- Body Position and Obstructive Sleep Apnea in Children: Sleep-Related Breathing DisordersDocument6 pagesBody Position and Obstructive Sleep Apnea in Children: Sleep-Related Breathing DisordersAnonymous 0Mnt71No ratings yet
- Jurnal 3 CDocument36 pagesJurnal 3 CCintya RambuNo ratings yet
- An Overview of Sleep and Circadian Dysfunction in Parkinson DiseaseDocument22 pagesAn Overview of Sleep and Circadian Dysfunction in Parkinson DiseaseOscar RodriguezNo ratings yet
- Learn. Mem.-2009-Jessberger-147-54llDocument9 pagesLearn. Mem.-2009-Jessberger-147-54llJean Pierre Chastre LuzaNo ratings yet
- Effects of Red Light On Sleep InertiaDocument13 pagesEffects of Red Light On Sleep InertiaMonica SaavedraNo ratings yet
- Glutamate Elicits Therapeutic Responses in Light-Induced Sleep-Deprived Zebrafish, Danio RerioDocument7 pagesGlutamate Elicits Therapeutic Responses in Light-Induced Sleep-Deprived Zebrafish, Danio RerioDr Sharique AliNo ratings yet
- The Relationship Between Sleep Problems and Neuropsychological FunctioningDocument7 pagesThe Relationship Between Sleep Problems and Neuropsychological FunctioningnoiNo ratings yet
- Elliott 2014Document5 pagesElliott 2014irnyirnyNo ratings yet
- 10 1093@sleep@26 2 117Document11 pages10 1093@sleep@26 2 117Intan AyueNo ratings yet
- Sleep Bruxism and Oromandibular Myoclonus in Rapid Eye Movement..., 2013Document7 pagesSleep Bruxism and Oromandibular Myoclonus in Rapid Eye Movement..., 2013Carlos Alberto Carranza LópezNo ratings yet
- Blume Et Al - 2018 - Standing Sentinel During Human SleepDocument11 pagesBlume Et Al - 2018 - Standing Sentinel During Human SleepChandan ShandilNo ratings yet
- Mecanismos Genéticos Que Median La Regulación Circadiana Del SueñoDocument10 pagesMecanismos Genéticos Que Median La Regulación Circadiana Del SueñoDiego Soto ChavezNo ratings yet
- 1-s2.0-S0920121122001504-mainDocument7 pages1-s2.0-S0920121122001504-mainKrisztián TóthNo ratings yet
- CitizannnDocument58 pagesCitizannnnajla munawwaraNo ratings yet
- EEG Normal Sleep - StatPearls - NCBI BookshelfDocument7 pagesEEG Normal Sleep - StatPearls - NCBI BookshelfChandan ShandilNo ratings yet
- Nihms 1729238Document39 pagesNihms 1729238mpssctsunamNo ratings yet
- Neurogeneses Hipocampo 2013Document9 pagesNeurogeneses Hipocampo 2013Denise GarciaNo ratings yet
- REFERENCIA 49 2021 MALSAGOVA NOS3 Glu298Asp (G894T)Document9 pagesREFERENCIA 49 2021 MALSAGOVA NOS3 Glu298Asp (G894T)Karen RoblesNo ratings yet
- REFERENCIA 30 2021 MALSAGOVA AGT rs699 (M235T)Document6 pagesREFERENCIA 30 2021 MALSAGOVA AGT rs699 (M235T)Karen RoblesNo ratings yet
- Referencia 38 2021 Malsagova Actn3 r577xDocument37 pagesReferencia 38 2021 Malsagova Actn3 r577xKaren RoblesNo ratings yet
- Referencia 35 2021 Malsagova Ace IdDocument5 pagesReferencia 35 2021 Malsagova Ace IdKaren RoblesNo ratings yet
- Zhu 19Document12 pagesZhu 19Karen RoblesNo ratings yet
- Activity and Exercise PatternDocument20 pagesActivity and Exercise PatternRaquel M. MendozaNo ratings yet
- Confined Space EntryDocument2 pagesConfined Space Entryaudit jogiNo ratings yet
- The Anatomy and Action of The Horse PDFDocument168 pagesThe Anatomy and Action of The Horse PDFstefm100% (1)
- Manuarl Instruction Stern Weber Dental UDocument84 pagesManuarl Instruction Stern Weber Dental UevilboarderNo ratings yet
- PgsgaDocument2 pagesPgsgaArsela Rindang AzhariNo ratings yet
- Examples of Blog WritingDocument7 pagesExamples of Blog WritingprimistartNo ratings yet
- Ebs429 Assignment1Document1 pageEbs429 Assignment1NNadiah IsaNo ratings yet
- Occupational Safety and Health STDDocument65 pagesOccupational Safety and Health STDKirk Corales100% (4)
- Clinical Teaching MethodsDocument29 pagesClinical Teaching MethodsDr. Jayesh Patidar100% (7)
- Care and Management of Turkey, Duck andDocument35 pagesCare and Management of Turkey, Duck andShealla Pis-oNo ratings yet
- Risk Assessment and Analysis For Sliding Gates: Analysis of Gate Structure To Be MovedDocument6 pagesRisk Assessment and Analysis For Sliding Gates: Analysis of Gate Structure To Be Movedsmart_onyx17No ratings yet
- Critical Thinking 9-16Document17 pagesCritical Thinking 9-16chelseeNo ratings yet
- Psych 2Document24 pagesPsych 2julia484838No ratings yet
- Philips Titration Protocol 2017Document12 pagesPhilips Titration Protocol 2017Sam WhiteNo ratings yet
- Responsible Parenthood 2Document20 pagesResponsible Parenthood 2jessicadimailigNo ratings yet
- Final MCN-RLE - Journal Reading - AlabastroDocument3 pagesFinal MCN-RLE - Journal Reading - AlabastroEthel May AlabastroNo ratings yet
- Official: Á51Ñ Antimicrobial Effectiveness TestingDocument3 pagesOfficial: Á51Ñ Antimicrobial Effectiveness TestingKaren Rojas LópezNo ratings yet
- Ringworm (Tinea Capitis)Document15 pagesRingworm (Tinea Capitis)farmasi_hmNo ratings yet
- Book Review Why Nations FailDocument11 pagesBook Review Why Nations FailCarloSangronesNo ratings yet
- IMNCIDocument13 pagesIMNCIJayalakshmiullasNo ratings yet
- Overview of The Pharmaceutical Market in MoroccoDocument4 pagesOverview of The Pharmaceutical Market in MoroccoMohnish Alka ThakkarNo ratings yet
- Lec. 4 Cell InjuryDocument6 pagesLec. 4 Cell InjuryMehdi MohammedNo ratings yet
- 2015 - Effects of Deep Pressure Stimulation On Physiological Arousal VAYU VEST With STRESSOR MORON TESTDocument5 pages2015 - Effects of Deep Pressure Stimulation On Physiological Arousal VAYU VEST With STRESSOR MORON TESTM Izzur MaulaNo ratings yet
- PEPA SummaryDocument6 pagesPEPA SummaryKhalid Masood GhaniNo ratings yet
- Dayton Letter To IBMDocument5 pagesDayton Letter To IBMMinnesota Public RadioNo ratings yet
- Labconco ManualDocument69 pagesLabconco ManualFredy Fernando Huarhua GarciaNo ratings yet
- Case History Sample 2 Case History Sample 2Document6 pagesCase History Sample 2 Case History Sample 2einsteinNo ratings yet
- Introduction To Internal Medicine - PPTMDocument30 pagesIntroduction To Internal Medicine - PPTMAddyNo ratings yet
- Cardiovascular and Respiratory Systems - Unit TestDocument7 pagesCardiovascular and Respiratory Systems - Unit Testapi-305647618No ratings yet




























































































Download as pdf or txt
You might also like
- F& C OSCE Notes - University of Manchester, WL GanDocument57 pagesF& C OSCE Notes - University of Manchester, WL GanWeh Loong Gan100% (7)
- Neurofibromin 1 Regulates Early Developmental Sleep in DrosophilaDocument9 pagesNeurofibromin 1 Regulates Early Developmental Sleep in DrosophilaFernandoNo ratings yet
- A. Basic Sleep Science V. Arousal and BehaviorDocument2 pagesA. Basic Sleep Science V. Arousal and BehaviorivanNo ratings yet
- SleepDocument9 pagesSleepLobnaNo ratings yet
- Benefits of Napping and An Extended Duration of Recovery Sleep On Alertness and Immune Cells After Acute Sleep RestrictionDocument9 pagesBenefits of Napping and An Extended Duration of Recovery Sleep On Alertness and Immune Cells After Acute Sleep RestrictionMatheus VilarinhoNo ratings yet
- 1 s2.0 S0896627313006533 MainDocument13 pages1 s2.0 S0896627313006533 Mainshriram1082883No ratings yet
- Cho 2013Document4 pagesCho 2013Kassandra Alfaro AlvarezNo ratings yet
- Sleepdisturbanceslinked Togeneticdisorders: Rimawati TedjasukmanaDocument10 pagesSleepdisturbanceslinked Togeneticdisorders: Rimawati TedjasukmanaLeilybadryaNo ratings yet
- Immanuel 2014Document4 pagesImmanuel 2014Mary Alarcon PalominoNo ratings yet
- Sleep Paralysis: June 2012Document8 pagesSleep Paralysis: June 2012renuka1011No ratings yet
- SCIENCE 2006 Detecting Awareness in The Vegetative StateDocument1 pageSCIENCE 2006 Detecting Awareness in The Vegetative StatepreghieraperlavitaNo ratings yet
- Creatine Supplementation Reduces Sleep Need and Homeostatic Sleep Pressure in RatsDocument9 pagesCreatine Supplementation Reduces Sleep Need and Homeostatic Sleep Pressure in RatswNo ratings yet
- Parasonias em REMDocument16 pagesParasonias em REMhcrislopes5617No ratings yet
- Sleep Medicine: Original ArticleDocument11 pagesSleep Medicine: Original ArticleaviyahnikmahNo ratings yet
- Are There Parallel Channels in The Vestibular Nerve?: Ellengene H. PetersonDocument8 pagesAre There Parallel Channels in The Vestibular Nerve?: Ellengene H. PetersonramopavelNo ratings yet
- Sleep Paralysis: June 2012Document8 pagesSleep Paralysis: June 2012edamwone04No ratings yet
- 1 s2.0 S1095643321002221 Main 1Document12 pages1 s2.0 S1095643321002221 Main 11603365No ratings yet
- Human Neurobiolog y Sleep Sleep DeprivatDocument6 pagesHuman Neurobiolog y Sleep Sleep Deprivatmelgarhoward90No ratings yet
- Neurostimulation-techniques-to-enhance-sleep-and-improve-cognition-in-agingDocument16 pagesNeurostimulation-techniques-to-enhance-sleep-and-improve-cognition-in-agingPrakash N KNo ratings yet
- 523 FullDocument3 pages523 FulldesckygamerNo ratings yet
- Sleep in Gerstmann-Straüssler-Scheinker Disease.Document5 pagesSleep in Gerstmann-Straüssler-Scheinker Disease.Lídia YoshiharaNo ratings yet
- Vision Research: David H. Mcdougal, Paul D. GamlinDocument16 pagesVision Research: David H. Mcdougal, Paul D. GamlinCristina GómezNo ratings yet
- Sleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerriDocument9 pagesSleep Medicine: Oliviero Bruni, Luana Novelli, Silvia Miano, Liborio Parrino, Mario Giovanni Terzano, Raffaele FerripsicosmosNo ratings yet
- The Importance of Astrocyte-Derived Purines in The Modulation of SleepDocument11 pagesThe Importance of Astrocyte-Derived Purines in The Modulation of Sleepapi-214563130No ratings yet
- Borbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Document7 pagesBorbely AA Et Wirz-Justice A (1982) Human Neurobiol 1 205-2-With-Cover-Page-V2Sebastian PereiraNo ratings yet
- Edwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitsDocument16 pagesEdwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitssbachorickNo ratings yet
- Short Sleep - Poor Sleep? A Polysomnographic Study in A Large Population Based Sample of WomenDocument7 pagesShort Sleep - Poor Sleep? A Polysomnographic Study in A Large Population Based Sample of Womenmahfuz emonNo ratings yet
- Homer1a Is A Core Brain Molecular Correlate of Sleep LossDocument6 pagesHomer1a Is A Core Brain Molecular Correlate of Sleep LosswestNo ratings yet
- Sleep and Wakefulness in Drosophila MelanogasterDocument7 pagesSleep and Wakefulness in Drosophila MelanogasterFernandoNo ratings yet
- Astrocito y SueñoDocument17 pagesAstrocito y SueñoLEISLY TATIANA OVIEDO GOMEZNo ratings yet
- Extended Wakefulness: Compromised Metabolics in and Degeneration of Locus Ceruleus NeuronsDocument14 pagesExtended Wakefulness: Compromised Metabolics in and Degeneration of Locus Ceruleus NeuronsRehaan FayazNo ratings yet
- Des Car GableDocument10 pagesDes Car GableDago CastilloNo ratings yet
- HHS Public Access: Impact of Sleep Disturbances On Neurodegeneration: Insight From Studies in Animal ModelsDocument25 pagesHHS Public Access: Impact of Sleep Disturbances On Neurodegeneration: Insight From Studies in Animal ModelsPawan MishraNo ratings yet
- A Role For Non-Rapid-Eye-Movement Sleep Homeostasis in Perceptual LearningDocument7 pagesA Role For Non-Rapid-Eye-Movement Sleep Homeostasis in Perceptual LearningPravin KumarNo ratings yet
- Sleep in Drosophila and Its ContextDocument19 pagesSleep in Drosophila and Its ContextFernandoNo ratings yet
- 2001 Atlas, Rules, and Recording Techniques For The Scoring of Cyclic Alternating Pattern (CAP) in Human SleepDocument17 pages2001 Atlas, Rules, and Recording Techniques For The Scoring of Cyclic Alternating Pattern (CAP) in Human SleepNHUNG Hoàng HuyềnNo ratings yet
- Epilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CookDocument11 pagesEpilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CooknurhikmahNo ratings yet
- Manipulations of Amyloid Precursor Protein Cleavage Disrupt The Circadian Clock in Aging DrosophilaDocument10 pagesManipulations of Amyloid Precursor Protein Cleavage Disrupt The Circadian Clock in Aging DrosophilaRan LinNo ratings yet
- Sleep 26 2 117Document10 pagesSleep 26 2 117ClaudiaNo ratings yet
- Nocturnal Light Exposure Impairs Affective Responses in A Wavelength-Dependent MannerDocument7 pagesNocturnal Light Exposure Impairs Affective Responses in A Wavelength-Dependent MannerLiana DimaNo ratings yet
- 1 s2.0 S0167876016304329 MainDocument1 page1 s2.0 S0167876016304329 Mainmincuandrada10No ratings yet
- Whitehead 2018 Emergence of Mature Cortical Activity in Wakefulness and Sleep in Healthy Preterm and Full Term InfantsDocument9 pagesWhitehead 2018 Emergence of Mature Cortical Activity in Wakefulness and Sleep in Healthy Preterm and Full Term InfantsC SNo ratings yet
- Baird NeuroImage 16 1120 2002 PDFDocument7 pagesBaird NeuroImage 16 1120 2002 PDFVezér RóbertNo ratings yet
- Body Position and Obstructive Sleep Apnea in Children: Sleep-Related Breathing DisordersDocument6 pagesBody Position and Obstructive Sleep Apnea in Children: Sleep-Related Breathing DisordersAnonymous 0Mnt71No ratings yet
- Jurnal 3 CDocument36 pagesJurnal 3 CCintya RambuNo ratings yet
- An Overview of Sleep and Circadian Dysfunction in Parkinson DiseaseDocument22 pagesAn Overview of Sleep and Circadian Dysfunction in Parkinson DiseaseOscar RodriguezNo ratings yet
- Learn. Mem.-2009-Jessberger-147-54llDocument9 pagesLearn. Mem.-2009-Jessberger-147-54llJean Pierre Chastre LuzaNo ratings yet
- Effects of Red Light On Sleep InertiaDocument13 pagesEffects of Red Light On Sleep InertiaMonica SaavedraNo ratings yet
- Glutamate Elicits Therapeutic Responses in Light-Induced Sleep-Deprived Zebrafish, Danio RerioDocument7 pagesGlutamate Elicits Therapeutic Responses in Light-Induced Sleep-Deprived Zebrafish, Danio RerioDr Sharique AliNo ratings yet
- The Relationship Between Sleep Problems and Neuropsychological FunctioningDocument7 pagesThe Relationship Between Sleep Problems and Neuropsychological FunctioningnoiNo ratings yet
- Elliott 2014Document5 pagesElliott 2014irnyirnyNo ratings yet
- 10 1093@sleep@26 2 117Document11 pages10 1093@sleep@26 2 117Intan AyueNo ratings yet
- Sleep Bruxism and Oromandibular Myoclonus in Rapid Eye Movement..., 2013Document7 pagesSleep Bruxism and Oromandibular Myoclonus in Rapid Eye Movement..., 2013Carlos Alberto Carranza LópezNo ratings yet
- Blume Et Al - 2018 - Standing Sentinel During Human SleepDocument11 pagesBlume Et Al - 2018 - Standing Sentinel During Human SleepChandan ShandilNo ratings yet
- Mecanismos Genéticos Que Median La Regulación Circadiana Del SueñoDocument10 pagesMecanismos Genéticos Que Median La Regulación Circadiana Del SueñoDiego Soto ChavezNo ratings yet
- 1-s2.0-S0920121122001504-mainDocument7 pages1-s2.0-S0920121122001504-mainKrisztián TóthNo ratings yet
- CitizannnDocument58 pagesCitizannnnajla munawwaraNo ratings yet
- EEG Normal Sleep - StatPearls - NCBI BookshelfDocument7 pagesEEG Normal Sleep - StatPearls - NCBI BookshelfChandan ShandilNo ratings yet
- Nihms 1729238Document39 pagesNihms 1729238mpssctsunamNo ratings yet
- Neurogeneses Hipocampo 2013Document9 pagesNeurogeneses Hipocampo 2013Denise GarciaNo ratings yet
- REFERENCIA 49 2021 MALSAGOVA NOS3 Glu298Asp (G894T)Document9 pagesREFERENCIA 49 2021 MALSAGOVA NOS3 Glu298Asp (G894T)Karen RoblesNo ratings yet
- REFERENCIA 30 2021 MALSAGOVA AGT rs699 (M235T)Document6 pagesREFERENCIA 30 2021 MALSAGOVA AGT rs699 (M235T)Karen RoblesNo ratings yet
- Referencia 38 2021 Malsagova Actn3 r577xDocument37 pagesReferencia 38 2021 Malsagova Actn3 r577xKaren RoblesNo ratings yet
- Referencia 35 2021 Malsagova Ace IdDocument5 pagesReferencia 35 2021 Malsagova Ace IdKaren RoblesNo ratings yet
- Zhu 19Document12 pagesZhu 19Karen RoblesNo ratings yet
- Activity and Exercise PatternDocument20 pagesActivity and Exercise PatternRaquel M. MendozaNo ratings yet
- Confined Space EntryDocument2 pagesConfined Space Entryaudit jogiNo ratings yet
- The Anatomy and Action of The Horse PDFDocument168 pagesThe Anatomy and Action of The Horse PDFstefm100% (1)
- Manuarl Instruction Stern Weber Dental UDocument84 pagesManuarl Instruction Stern Weber Dental UevilboarderNo ratings yet
- PgsgaDocument2 pagesPgsgaArsela Rindang AzhariNo ratings yet
- Examples of Blog WritingDocument7 pagesExamples of Blog WritingprimistartNo ratings yet
- Ebs429 Assignment1Document1 pageEbs429 Assignment1NNadiah IsaNo ratings yet
- Occupational Safety and Health STDDocument65 pagesOccupational Safety and Health STDKirk Corales100% (4)
- Clinical Teaching MethodsDocument29 pagesClinical Teaching MethodsDr. Jayesh Patidar100% (7)
- Care and Management of Turkey, Duck andDocument35 pagesCare and Management of Turkey, Duck andShealla Pis-oNo ratings yet
- Risk Assessment and Analysis For Sliding Gates: Analysis of Gate Structure To Be MovedDocument6 pagesRisk Assessment and Analysis For Sliding Gates: Analysis of Gate Structure To Be Movedsmart_onyx17No ratings yet
- Critical Thinking 9-16Document17 pagesCritical Thinking 9-16chelseeNo ratings yet
- Psych 2Document24 pagesPsych 2julia484838No ratings yet
- Philips Titration Protocol 2017Document12 pagesPhilips Titration Protocol 2017Sam WhiteNo ratings yet
- Responsible Parenthood 2Document20 pagesResponsible Parenthood 2jessicadimailigNo ratings yet
- Final MCN-RLE - Journal Reading - AlabastroDocument3 pagesFinal MCN-RLE - Journal Reading - AlabastroEthel May AlabastroNo ratings yet
- Official: Á51Ñ Antimicrobial Effectiveness TestingDocument3 pagesOfficial: Á51Ñ Antimicrobial Effectiveness TestingKaren Rojas LópezNo ratings yet
- Ringworm (Tinea Capitis)Document15 pagesRingworm (Tinea Capitis)farmasi_hmNo ratings yet
- Book Review Why Nations FailDocument11 pagesBook Review Why Nations FailCarloSangronesNo ratings yet
- IMNCIDocument13 pagesIMNCIJayalakshmiullasNo ratings yet
- Overview of The Pharmaceutical Market in MoroccoDocument4 pagesOverview of The Pharmaceutical Market in MoroccoMohnish Alka ThakkarNo ratings yet
- Lec. 4 Cell InjuryDocument6 pagesLec. 4 Cell InjuryMehdi MohammedNo ratings yet
- 2015 - Effects of Deep Pressure Stimulation On Physiological Arousal VAYU VEST With STRESSOR MORON TESTDocument5 pages2015 - Effects of Deep Pressure Stimulation On Physiological Arousal VAYU VEST With STRESSOR MORON TESTM Izzur MaulaNo ratings yet
- PEPA SummaryDocument6 pagesPEPA SummaryKhalid Masood GhaniNo ratings yet
- Dayton Letter To IBMDocument5 pagesDayton Letter To IBMMinnesota Public RadioNo ratings yet
- Labconco ManualDocument69 pagesLabconco ManualFredy Fernando Huarhua GarciaNo ratings yet
- Case History Sample 2 Case History Sample 2Document6 pagesCase History Sample 2 Case History Sample 2einsteinNo ratings yet
- Introduction To Internal Medicine - PPTMDocument30 pagesIntroduction To Internal Medicine - PPTMAddyNo ratings yet
- Cardiovascular and Respiratory Systems - Unit TestDocument7 pagesCardiovascular and Respiratory Systems - Unit Testapi-305647618No ratings yet