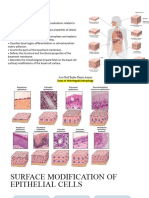
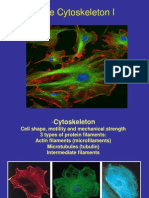
Download as pdf or txt
You might also like
- Chapter 9 Homework PDFDocument3 pagesChapter 9 Homework PDFRhonda JacobsNo ratings yet
- Microfilament: in Vitro Self-AssemblyDocument7 pagesMicrofilament: in Vitro Self-AssemblyABHISHEK RATHODNo ratings yet
- Cytology CytoskeletonDocument48 pagesCytology CytoskeletonMitzel Sapalo0% (1)
- MicrofilamentsDocument4 pagesMicrofilamentsSujan maharjanNo ratings yet
- Chapter 16 SummaryDocument11 pagesChapter 16 SummaryCharlotteNo ratings yet
- Cell Locomotion PDFDocument8 pagesCell Locomotion PDFmanoj_rkl_07No ratings yet
- 14 The CytoskeletonDocument7 pages14 The Cytoskeletonsatriamanullang880No ratings yet
- C YTOSKELETONDocument44 pagesC YTOSKELETONFatma Nadjah MandaNo ratings yet
- Cytoskeleton Centrosome and CentrioleDocument22 pagesCytoskeleton Centrosome and CentriolekdbuereNo ratings yet
- Cy To SkeletonDocument21 pagesCy To SkeletonJisna JaisonNo ratings yet
- LO8Document1 pageLO8Yo TuNo ratings yet
- 2 Basal Lateral Apical Modifications and Connective TissueDocument84 pages2 Basal Lateral Apical Modifications and Connective TissueDeniz asmazNo ratings yet
- Actin Dynamics in Cell MigrationDocument13 pagesActin Dynamics in Cell Migration王麦No ratings yet
- Cell Morphology and Motility: Peter Takizawa Cell BiologyDocument41 pagesCell Morphology and Motility: Peter Takizawa Cell BiologyMasresha AhmedNo ratings yet
- Cytoskeleton DADocument35 pagesCytoskeleton DAAnonymous NVBCqKWHNo ratings yet
- Inigo.j 01109151013Document27 pagesInigo.j 01109151013inigoNo ratings yet
- Actin Microfilaments Study GuideDocument2 pagesActin Microfilaments Study GuideBlaNo ratings yet
- Mammalian Histology AssignmentDocument9 pagesMammalian Histology AssignmentSana Sultana100% (1)
- Introduction To Molecular Motor Proteins: Part 1 Lecture NotesDocument12 pagesIntroduction To Molecular Motor Proteins: Part 1 Lecture Noteskinza888No ratings yet
- Analyzing Circulating Tumor Cells One at A Time: HighlightsDocument22 pagesAnalyzing Circulating Tumor Cells One at A Time: Highlightsian merisNo ratings yet
- Lecture 3 - Chapter 8-Cytoskeleton ADocument75 pagesLecture 3 - Chapter 8-Cytoskeleton AKw Chan33% (3)
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- Traffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlDocument8 pagesTraffic - 2004 - Etienne Manneville - Actin and Microtubules in Cell Motility Which One Is in ControlPinaki NayakNo ratings yet
- Structure of Microtubules Amd ActinsDocument19 pagesStructure of Microtubules Amd Actinspreetakaran12No ratings yet
- February 27, 2012 - Chapter 7: Inside The CellDocument4 pagesFebruary 27, 2012 - Chapter 7: Inside The CellVincent Troy SantosNo ratings yet
- Cell Structure, Organization, Bacteria and Archaea: Defining StatementDocument13 pagesCell Structure, Organization, Bacteria and Archaea: Defining StatementRoger John PeñasNo ratings yet
- A New Link Between The Retrograde Actin Flow and FDocument11 pagesA New Link Between The Retrograde Actin Flow and FPinaki NayakNo ratings yet
- Biology Lecture 3: The Cell 1: 1.describe The Structure and Function of The Components of The Endomembrane SystemDocument3 pagesBiology Lecture 3: The Cell 1: 1.describe The Structure and Function of The Components of The Endomembrane Systemphuc thienNo ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- Centro, MF. Module - 6.Document9 pagesCentro, MF. Module - 6.jessa marie marevilesNo ratings yet
- Motor ProteinsDocument9 pagesMotor Proteinsazazel666No ratings yet
- CHAP 2. CSH TẾ BÀO - 232Document100 pagesCHAP 2. CSH TẾ BÀO - 232Nguyễn Anh QuangNo ratings yet
- 2cytology Cytoskel Receptor Transp (3) 1Document43 pages2cytology Cytoskel Receptor Transp (3) 1m.mansoor3377No ratings yet
- Bio 201 Part 2Document45 pagesBio 201 Part 2Vivian WuNo ratings yet
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- Chapter 16 - The Cytoskeleton - 112612Document56 pagesChapter 16 - The Cytoskeleton - 112612Sohan rautNo ratings yet
- Chapter 2 NeuronDocument49 pagesChapter 2 NeuronAfNo ratings yet
- Cell Structures and FunctionsDocument5 pagesCell Structures and Functions9pjh6pzxmsNo ratings yet
- The CytoskeletonDocument63 pagesThe CytoskeletonAmara TargaryenNo ratings yet
- Cell Signaling 5 LessonDocument66 pagesCell Signaling 5 LessonPrathamesh KulkarniNo ratings yet
- BY2201 EmmaC CytoskeletonIandIIDocument62 pagesBY2201 EmmaC CytoskeletonIandIIRebecca ReillyNo ratings yet
- Cytoskeletal SystemDocument28 pagesCytoskeletal Systemswarnaprabhadas07No ratings yet
- Cell Adhesion, Cell Movement and Extracellular MatrixDocument17 pagesCell Adhesion, Cell Movement and Extracellular MatrixAD-MQNo ratings yet
- Microfilament Actin 1Document19 pagesMicrofilament Actin 1DwiRaniPrihandiniNo ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- Chapter 2 - NotesDocument22 pagesChapter 2 - NotesanusoumyaNo ratings yet
- CELLMOLDocument3 pagesCELLMOLJhon Rey LagosNo ratings yet
- Cytoskeleton: Jetro P. MagnayonDocument66 pagesCytoskeleton: Jetro P. MagnayonChristian Santiago100% (1)
- 4.1 CytoskeletonDocument28 pages4.1 Cytoskeletonedabzan234No ratings yet
- Lecture 32 - Actin Filaments - Post-LectureDocument47 pagesLecture 32 - Actin Filaments - Post-LectureMuhit ChowdhuryNo ratings yet
- Capping Protein Regulators Fine-Tune Actin Assembly DynamicsDocument29 pagesCapping Protein Regulators Fine-Tune Actin Assembly DynamicsGABRIELA GERALDINE SOTO ACEVEDONo ratings yet
- Organelles Working TogetherDocument4 pagesOrganelles Working TogetherVilma Malaya VisarioNo ratings yet
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Epithelial Tissues REVIEWERDocument16 pagesEpithelial Tissues REVIEWERClyde BaltazarNo ratings yet
- CytoskeletonDocument65 pagesCytoskeletonSapana SubediNo ratings yet
- Histology 3 - Cytoplasm 4 & NucleusDocument8 pagesHistology 3 - Cytoplasm 4 & NucleusShariffa KhadijaNo ratings yet
- Chapter Overview: Subject Name The Cell Topic 1 Week 1Document9 pagesChapter Overview: Subject Name The Cell Topic 1 Week 1VeiliLookNo ratings yet
- Amino Acids and ProteinsDocument2 pagesAmino Acids and ProteinsyejiNo ratings yet
- Enumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonDocument10 pagesEnumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonTathagata ChakravarttyNo ratings yet
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- BI309 Ryan Carroll Lab 1 For HADocument4 pagesBI309 Ryan Carroll Lab 1 For HARyan CarrollNo ratings yet
- BI309 Lab 3 Ryan CarrollDocument4 pagesBI309 Lab 3 Ryan CarrollRyan CarrollNo ratings yet
- Assignment 2 - Noncovalent Bonds in Proteins Ryan Carroll, 19496206Document2 pagesAssignment 2 - Noncovalent Bonds in Proteins Ryan Carroll, 19496206Ryan CarrollNo ratings yet
- Assignment 2Document2 pagesAssignment 2Ryan CarrollNo ratings yet
- The Birth and Death of Proteins: Some Key ConceptsDocument2 pagesThe Birth and Death of Proteins: Some Key ConceptsLuke ShantiNo ratings yet
- Biological Tools TechniquesDocument10 pagesBiological Tools Techniquesayushi75086No ratings yet
- Nucleus - Morphology and FunctionsDocument25 pagesNucleus - Morphology and FunctionsVytheeshwaran Vedagiri96% (24)
- Transcription ProkaryoticDocument30 pagesTranscription ProkaryoticDibya Jyoti ParidaNo ratings yet
- Assignment Question Db014 (2023 - 2024)Document3 pagesAssignment Question Db014 (2023 - 2024)yuni adnanNo ratings yet
- DNA Fingerprinting PowerpointDocument23 pagesDNA Fingerprinting PowerpointqonitaNo ratings yet
- Test Bank For Biochemistry A Short Course Third EditionDocument10 pagesTest Bank For Biochemistry A Short Course Third EditionJoshuaLeeegfrq100% (50)
- A Project On Protein Synthesis and ProcessingDocument25 pagesA Project On Protein Synthesis and ProcessingAnkita JalanNo ratings yet
- HIV Drug Resistance Training: Molecular Lab Set-Up and WorkflowDocument31 pagesHIV Drug Resistance Training: Molecular Lab Set-Up and WorkflowYantek Btklpp ManadoNo ratings yet
- Bioinformatics Scripting: Submitted To: Dr. Khuram ShahzadDocument4 pagesBioinformatics Scripting: Submitted To: Dr. Khuram ShahzadSidra chaudharyNo ratings yet
- Karolin Luger - Nucleosomes: Structure and FunctionDocument8 pagesKarolin Luger - Nucleosomes: Structure and FunctionDopameNo ratings yet
- Transcription and TranslationDocument27 pagesTranscription and Translationᴅᴇᴀᴅ ᴋɪʟʟᴇʀNo ratings yet
- Tecalco Cruz 2017 R3Document10 pagesTecalco Cruz 2017 R3Valentina Conde AndradeNo ratings yet
- DNA Sequencing: Chemical Method/ Maxam Gilbert MethodDocument19 pagesDNA Sequencing: Chemical Method/ Maxam Gilbert MethodZain YaqoobNo ratings yet
- Spirochaeta SP L21-RPul-D2 GFFDocument158 pagesSpirochaeta SP L21-RPul-D2 GFFMARCO ANTONIONo ratings yet
- Cytoskeleton 13.4.20Document39 pagesCytoskeleton 13.4.20NikhilNo ratings yet
- Gene Structure Lecture 2Document36 pagesGene Structure Lecture 2AlexTsuiNo ratings yet
- TRP Operon PresentationDocument27 pagesTRP Operon PresentationPradeep SinghNo ratings yet
- Blood Clotting MechanismDocument19 pagesBlood Clotting Mechanismchinecheremstephanie2022No ratings yet
- and Differentiation Markers in THP1: CaggtcaacctattggtcaagccDocument4 pagesand Differentiation Markers in THP1: CaggtcaacctattggtcaagccLara AiassaNo ratings yet
- Mitochondrial DNA Protocol - 2016Document233 pagesMitochondrial DNA Protocol - 2016Sivapong Vsmu100% (1)
- AP Bio U1Document92 pagesAP Bio U1朱國修No ratings yet
- Dna ReportDocument21 pagesDna Report168-Shivank DwivediNo ratings yet
- Genetic Analysis An Integrated Approach 2Nd Edition Sanders Test Bank Full Chapter PDFDocument35 pagesGenetic Analysis An Integrated Approach 2Nd Edition Sanders Test Bank Full Chapter PDFallison.young656100% (20)
- SOL Review Biochemistry and WaterDocument1 pageSOL Review Biochemistry and WaterAlayna SheltonNo ratings yet
- AP Biology Free Response On Protein SynthesisDocument7 pagesAP Biology Free Response On Protein Synthesisafktgvllsgajfw100% (2)
- Transcription Practice WorksheetDocument3 pagesTranscription Practice Worksheetbrimej4513No ratings yet
- Rticle: Structural and Functional Analysis of Cyclin D1 Reveals p27 and Substrate Inhibitor Binding RequirementsDocument14 pagesRticle: Structural and Functional Analysis of Cyclin D1 Reveals p27 and Substrate Inhibitor Binding RequirementsSamyabrata BhaduriNo ratings yet
- Wikipedia - Readings On Life Evolutionary History V2Document3,688 pagesWikipedia - Readings On Life Evolutionary History V2tariqamin5978No ratings yet