
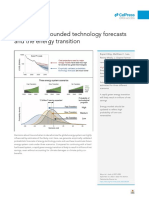
1 s2.0 S0960982213003485 Main
1 s2.0 S0960982213003485 Main
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Natural Selection Lab-Phet SimulationDocument6 pagesNatural Selection Lab-Phet Simulationapi-315483634No ratings yet
- Genetic Programming Seminar ReportDocument39 pagesGenetic Programming Seminar ReportProloy GhoshNo ratings yet
- Analyses of Cross Sectional Data To Link The PEG With The Patie - 2022 - The JouDocument8 pagesAnalyses of Cross Sectional Data To Link The PEG With The Patie - 2022 - The JouFeyzullah TokgözNo ratings yet
- 1 s2.0 S1526590022003662 MainDocument10 pages1 s2.0 S1526590022003662 MainFeyzullah TokgözNo ratings yet
- 1 s2.0 S254243512200410X MainDocument27 pages1 s2.0 S254243512200410X MainFeyzullah TokgözNo ratings yet
- 1 s2.0 S096098221300362X MainDocument2 pages1 s2.0 S096098221300362X MainFeyzullah TokgözNo ratings yet
- 1 s2.0 S1578219020300792 MainDocument2 pages1 s2.0 S1578219020300792 MainFeyzullah TokgözNo ratings yet
- Denkl emDocument27 pagesDenkl emFeyzullah TokgözNo ratings yet
- Reversal DistanceDocument5 pagesReversal Distanceviveksahu87No ratings yet
- CH 16 WorkbookDocument10 pagesCH 16 Workbookapi-237586441No ratings yet
- Lightning Critical Flashover Voltage of High Voltage Insulators: Laboratory Measurements and CalculationsDocument9 pagesLightning Critical Flashover Voltage of High Voltage Insulators: Laboratory Measurements and CalculationsLeonardo MendozaNo ratings yet
- Microevolutionary Forcess PDFDocument42 pagesMicroevolutionary Forcess PDFdewi utariNo ratings yet
- Design Optimization of Electrical Transformer Using Genetic Algorithm Phaengkieo2014Document5 pagesDesign Optimization of Electrical Transformer Using Genetic Algorithm Phaengkieo2014Beurus SamaNo ratings yet
- Module 3 GEE 4Document22 pagesModule 3 GEE 4abegail arponNo ratings yet
- 1 The Evolution of Alternative Reproductive Tactics: Concepts and QuestionsDocument21 pages1 The Evolution of Alternative Reproductive Tactics: Concepts and QuestionsAlexandra ElenaNo ratings yet
- GA Toolbox in MatlabDocument105 pagesGA Toolbox in MatlabSoy GuajiroNo ratings yet
- Exit Mcqs.-1Document180 pagesExit Mcqs.-1Muhammad Yasin Sabir0% (1)
- Practice 1 So LNDocument5 pagesPractice 1 So LNAdesh Nidhi TiwaryNo ratings yet
- Ecology Unit Objective ChecklistDocument3 pagesEcology Unit Objective Checklistapi-281248095No ratings yet
- AnswersDocument19 pagesAnswersSherlock Wesley ConanNo ratings yet
- The Evolution of Biological Information How Evolution Creates Complexity From Viruses To Brains Christoph Adami Full Chapter PDFDocument69 pagesThe Evolution of Biological Information How Evolution Creates Complexity From Viruses To Brains Christoph Adami Full Chapter PDFhirukusaifee100% (3)
- Automation of Exam Hall Allotment and Seating Arrangement IJERTV10IS060193Document6 pagesAutomation of Exam Hall Allotment and Seating Arrangement IJERTV10IS060193Aarush VermaNo ratings yet
- Cia Assgn1 201516Document6 pagesCia Assgn1 201516Diego Jose Palaquibay IngaNo ratings yet
- Foundations of Learning and Adaptive Systems: Evolutionary AlgorithmsDocument26 pagesFoundations of Learning and Adaptive Systems: Evolutionary AlgorithmsKelvin KarisNo ratings yet
- Study Guide Key CH - 23 - MicroevolutionDocument5 pagesStudy Guide Key CH - 23 - Microevolutionrulz65100% (1)
- 3-The Newest Synthesis - Understanding The Interplay of Evolutionary and Ecologycal Dynamicsw - Schoener - Science-2011Document5 pages3-The Newest Synthesis - Understanding The Interplay of Evolutionary and Ecologycal Dynamicsw - Schoener - Science-2011Uriel RodríguezNo ratings yet
- Genetic Algorithms and Game Theory For Airport Departure Decision Making: Gedman and CodmanDocument13 pagesGenetic Algorithms and Game Theory For Airport Departure Decision Making: Gedman and CodmanAhmedNo ratings yet
- Evo101 - 03 - Mechanisms - The Process of Evolution - UEDocument34 pagesEvo101 - 03 - Mechanisms - The Process of Evolution - UEJanice LerioratoNo ratings yet
- Friendship and The Banker's ParadoxDocument26 pagesFriendship and The Banker's ParadoxozzycattNo ratings yet
- Auto DockDocument13 pagesAuto DockKarthick YuvrajNo ratings yet
- Chapter 3 - Lecturer 2 Other Nature Inspired Optimization TechniquesDocument32 pagesChapter 3 - Lecturer 2 Other Nature Inspired Optimization TechniquesMK JAYANTHI KANNANNo ratings yet
- Principles of Social Psychology - AttributedDocument752 pagesPrinciples of Social Psychology - AttributedAlfonso J Sintjago100% (1)
- As Distintas Faces Do ComportamentoDocument426 pagesAs Distintas Faces Do Comportamentobarbosa03100% (1)
- TautologyDocument5 pagesTautologySuhaila AlDunayNo ratings yet
- Genetic AlgorithmDocument57 pagesGenetic AlgorithmRushikesh DandagwhalNo ratings yet










































































Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5823)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (541)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (823)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Natural Selection Lab-Phet SimulationDocument6 pagesNatural Selection Lab-Phet Simulationapi-315483634No ratings yet
- Genetic Programming Seminar ReportDocument39 pagesGenetic Programming Seminar ReportProloy GhoshNo ratings yet
- Analyses of Cross Sectional Data To Link The PEG With The Patie - 2022 - The JouDocument8 pagesAnalyses of Cross Sectional Data To Link The PEG With The Patie - 2022 - The JouFeyzullah TokgözNo ratings yet
- 1 s2.0 S1526590022003662 MainDocument10 pages1 s2.0 S1526590022003662 MainFeyzullah TokgözNo ratings yet
- 1 s2.0 S254243512200410X MainDocument27 pages1 s2.0 S254243512200410X MainFeyzullah TokgözNo ratings yet
- 1 s2.0 S096098221300362X MainDocument2 pages1 s2.0 S096098221300362X MainFeyzullah TokgözNo ratings yet
- 1 s2.0 S1578219020300792 MainDocument2 pages1 s2.0 S1578219020300792 MainFeyzullah TokgözNo ratings yet
- Denkl emDocument27 pagesDenkl emFeyzullah TokgözNo ratings yet
- Reversal DistanceDocument5 pagesReversal Distanceviveksahu87No ratings yet
- CH 16 WorkbookDocument10 pagesCH 16 Workbookapi-237586441No ratings yet
- Lightning Critical Flashover Voltage of High Voltage Insulators: Laboratory Measurements and CalculationsDocument9 pagesLightning Critical Flashover Voltage of High Voltage Insulators: Laboratory Measurements and CalculationsLeonardo MendozaNo ratings yet
- Microevolutionary Forcess PDFDocument42 pagesMicroevolutionary Forcess PDFdewi utariNo ratings yet
- Design Optimization of Electrical Transformer Using Genetic Algorithm Phaengkieo2014Document5 pagesDesign Optimization of Electrical Transformer Using Genetic Algorithm Phaengkieo2014Beurus SamaNo ratings yet
- Module 3 GEE 4Document22 pagesModule 3 GEE 4abegail arponNo ratings yet
- 1 The Evolution of Alternative Reproductive Tactics: Concepts and QuestionsDocument21 pages1 The Evolution of Alternative Reproductive Tactics: Concepts and QuestionsAlexandra ElenaNo ratings yet
- GA Toolbox in MatlabDocument105 pagesGA Toolbox in MatlabSoy GuajiroNo ratings yet
- Exit Mcqs.-1Document180 pagesExit Mcqs.-1Muhammad Yasin Sabir0% (1)
- Practice 1 So LNDocument5 pagesPractice 1 So LNAdesh Nidhi TiwaryNo ratings yet
- Ecology Unit Objective ChecklistDocument3 pagesEcology Unit Objective Checklistapi-281248095No ratings yet
- AnswersDocument19 pagesAnswersSherlock Wesley ConanNo ratings yet
- The Evolution of Biological Information How Evolution Creates Complexity From Viruses To Brains Christoph Adami Full Chapter PDFDocument69 pagesThe Evolution of Biological Information How Evolution Creates Complexity From Viruses To Brains Christoph Adami Full Chapter PDFhirukusaifee100% (3)
- Automation of Exam Hall Allotment and Seating Arrangement IJERTV10IS060193Document6 pagesAutomation of Exam Hall Allotment and Seating Arrangement IJERTV10IS060193Aarush VermaNo ratings yet
- Cia Assgn1 201516Document6 pagesCia Assgn1 201516Diego Jose Palaquibay IngaNo ratings yet
- Foundations of Learning and Adaptive Systems: Evolutionary AlgorithmsDocument26 pagesFoundations of Learning and Adaptive Systems: Evolutionary AlgorithmsKelvin KarisNo ratings yet
- Study Guide Key CH - 23 - MicroevolutionDocument5 pagesStudy Guide Key CH - 23 - Microevolutionrulz65100% (1)
- 3-The Newest Synthesis - Understanding The Interplay of Evolutionary and Ecologycal Dynamicsw - Schoener - Science-2011Document5 pages3-The Newest Synthesis - Understanding The Interplay of Evolutionary and Ecologycal Dynamicsw - Schoener - Science-2011Uriel RodríguezNo ratings yet
- Genetic Algorithms and Game Theory For Airport Departure Decision Making: Gedman and CodmanDocument13 pagesGenetic Algorithms and Game Theory For Airport Departure Decision Making: Gedman and CodmanAhmedNo ratings yet
- Evo101 - 03 - Mechanisms - The Process of Evolution - UEDocument34 pagesEvo101 - 03 - Mechanisms - The Process of Evolution - UEJanice LerioratoNo ratings yet
- Friendship and The Banker's ParadoxDocument26 pagesFriendship and The Banker's ParadoxozzycattNo ratings yet
- Auto DockDocument13 pagesAuto DockKarthick YuvrajNo ratings yet
- Chapter 3 - Lecturer 2 Other Nature Inspired Optimization TechniquesDocument32 pagesChapter 3 - Lecturer 2 Other Nature Inspired Optimization TechniquesMK JAYANTHI KANNANNo ratings yet
- Principles of Social Psychology - AttributedDocument752 pagesPrinciples of Social Psychology - AttributedAlfonso J Sintjago100% (1)
- As Distintas Faces Do ComportamentoDocument426 pagesAs Distintas Faces Do Comportamentobarbosa03100% (1)
- TautologyDocument5 pagesTautologySuhaila AlDunayNo ratings yet
- Genetic AlgorithmDocument57 pagesGenetic AlgorithmRushikesh DandagwhalNo ratings yet