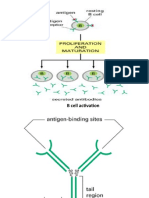
Download as pdf or txt
You might also like
- Nesters Microbiology A Human Perspective 10Th Edition Anderson Full ChapterDocument67 pagesNesters Microbiology A Human Perspective 10Th Edition Anderson Full Chapterjonathan.smith167100% (12)
- Janeways Immunobiology 9Th Edition Murphy Test Bank Full Chapter PDFDocument44 pagesJaneways Immunobiology 9Th Edition Murphy Test Bank Full Chapter PDFJamesWellsoeic100% (12)
- T CellDocument36 pagesT CellSurja DasNo ratings yet
- L 4 Cellular Component of The Immune SystemDocument11 pagesL 4 Cellular Component of The Immune SystemaugustongoviNo ratings yet
- Cells and Organs of The Immune SystemDocument26 pagesCells and Organs of The Immune SystemMithila BanikNo ratings yet
- محاضرةة الانسجةDocument17 pagesمحاضرةة الانسجةAymen MouradNo ratings yet
- Immuno 1 Unit I SemDocument5 pagesImmuno 1 Unit I Sempriyanshu guptaNo ratings yet
- Histology BloodDocument6 pagesHistology BloodNeneng Cutin GanzonNo ratings yet
- White Blood CellDocument10 pagesWhite Blood CellsujithasNo ratings yet
- AnatomyDocument2 pagesAnatomyIsaac FrankNo ratings yet
- Immuno PathologyDocument28 pagesImmuno PathologyIdrissa John Sebeh ContehNo ratings yet
- Cells of ImmunityDocument3 pagesCells of ImmunityLenc AbaloNo ratings yet
- Chapter Three: The Lymphoid SystemDocument4 pagesChapter Three: The Lymphoid SystemaiaiNo ratings yet
- Components of Immune SystemsDocument73 pagesComponents of Immune SystemsMaria Nadeem, BS Home Economics Student, UoPNo ratings yet
- Introduction of ImmunologyDocument33 pagesIntroduction of ImmunologyMenard Paisley MarchalNo ratings yet
- What Are The Main Cells of The Immune System?Document5 pagesWhat Are The Main Cells of The Immune System?Idrissa John Sebeh ContehNo ratings yet
- Immune System by Asif PresentationDocument35 pagesImmune System by Asif PresentationHassan AsifNo ratings yet
- Part 2 Basics of ImmunologyDocument6 pagesPart 2 Basics of Immunologyabc abcNo ratings yet
- Chap. RBC N WBC GoodDocument33 pagesChap. RBC N WBC GoodMahima SinhaNo ratings yet
- Biology - Cells and Anatomy of The Immune SystemDocument9 pagesBiology - Cells and Anatomy of The Immune SystemFelix LupulescuNo ratings yet
- Chapter 2 Cells and Organs of The Immune System PDFDocument61 pagesChapter 2 Cells and Organs of The Immune System PDFanaraudhatulNo ratings yet
- Lymphocytes and MonocytesDocument7 pagesLymphocytes and MonocytesSarfraz Hussain MalikNo ratings yet
- Organs of The Immune SystemDocument19 pagesOrgans of The Immune SystemprabuNo ratings yet
- Rajiv Gandhi University of Technology and Science School of Medicine MD1Document7 pagesRajiv Gandhi University of Technology and Science School of Medicine MD1Devika KhanNo ratings yet
- About LymphomaDocument17 pagesAbout LymphomaSelviNo ratings yet
- Basic Concepts in ImmunologyDocument56 pagesBasic Concepts in ImmunologyDedy SambahtraNo ratings yet
- The Cells of The Immune System: Aditya Gupta & Alok Singh S.Y.Bsc (Microbiology) V.E.S. CollegeDocument22 pagesThe Cells of The Immune System: Aditya Gupta & Alok Singh S.Y.Bsc (Microbiology) V.E.S. CollegevesascNo ratings yet
- Lecture - Cells Tissues and Organs - ImmunologyDocument29 pagesLecture - Cells Tissues and Organs - Immunologyrockyrawat01012003No ratings yet
- Topic 1,2,3 y 4Document15 pagesTopic 1,2,3 y 4Olaya De Dios HuertaNo ratings yet
- Immunology NotesDocument37 pagesImmunology NotesSanthosh Kalash100% (2)
- Irwansyah Nur Oktafian - 180342618025 - Immunology TasksDocument5 pagesIrwansyah Nur Oktafian - 180342618025 - Immunology TasksIRWANSYAH NURNo ratings yet
- Immune System Docs 2019Document13 pagesImmune System Docs 2019David DavidNo ratings yet
- Basic Immunology PDFDocument6 pagesBasic Immunology PDFAnastasia ENo ratings yet
- White Blood Cells - Components of Lymphatic SystemDocument26 pagesWhite Blood Cells - Components of Lymphatic Systemislamicmedia188No ratings yet
- 2.cells of Immune System Notes PowerpointDocument34 pages2.cells of Immune System Notes Powerpointt4ths999No ratings yet
- Lymphocytes - 12Document19 pagesLymphocytes - 12Nikhil BijuNo ratings yet
- Immunology: Dr. A.K.M. Akbar KabirDocument29 pagesImmunology: Dr. A.K.M. Akbar KabirRakib's exploration worldNo ratings yet
- Organs of Immune SystemDocument31 pagesOrgans of Immune Systemspourush100% (2)
- Hematopoiesis and Cells of The Immune SystemDocument19 pagesHematopoiesis and Cells of The Immune SystemCGM BF OFFICENo ratings yet
- Antibody 1Document19 pagesAntibody 1Silas AnkuNo ratings yet
- Hiv & AdisDocument60 pagesHiv & Adisomarism1960No ratings yet
- Cells of The Immune SystemDocument38 pagesCells of The Immune SystemprabuNo ratings yet
- Chapter 2 Basicc Immunology Ppts DZ 2010Document85 pagesChapter 2 Basicc Immunology Ppts DZ 2010TofikNo ratings yet
- A Simple Guide to the Blood Cells, Related Diseases And Use in Disease DiagnosisFrom EverandA Simple Guide to the Blood Cells, Related Diseases And Use in Disease DiagnosisNo ratings yet
- იმუნოს ზეპირი 1 1Document65 pagesიმუნოს ზეპირი 1 1Levani KartvelishviliNo ratings yet
- Копия Diseases - of - Immunity.-2021. - копияDocument77 pagesКопия Diseases - of - Immunity.-2021. - копияManav VyasNo ratings yet
- Bahan Kuliah Id-1-2020Document63 pagesBahan Kuliah Id-1-2020Andi SaputraNo ratings yet
- Fuctions of WBCsDocument3 pagesFuctions of WBCsTahir DocNo ratings yet
- Chapter 1immunology OverviewDocument37 pagesChapter 1immunology OverviewSania HairahNo ratings yet
- Chapter 2Document8 pagesChapter 2Zaheer AhmedNo ratings yet
- White Blood Cell (Leucocyte)Document77 pagesWhite Blood Cell (Leucocyte)Dr.Gomathi sivakumarNo ratings yet
- Immunology Overview: Armond S. Goldman Bellur S. PrabhakarDocument45 pagesImmunology Overview: Armond S. Goldman Bellur S. PrabhakarIoana AsziaNo ratings yet
- Course 11 - ImunochemistryDocument31 pagesCourse 11 - ImunochemistryPopa NicuNo ratings yet
- Guia de Estudios PCFDocument7 pagesGuia de Estudios PCFOmar AguilarNo ratings yet
- Dendritic Cell BiologyDocument12 pagesDendritic Cell BiologyHelena QuintNo ratings yet
- Immunology Presentation 2.0Document10 pagesImmunology Presentation 2.0Emmanuel KwakunakaNo ratings yet
- Name: Abby Yong I.D.: 14WAR03484 Group: RBS2 Group 2 Date: 30 September 2015Document10 pagesName: Abby Yong I.D.: 14WAR03484 Group: RBS2 Group 2 Date: 30 September 2015Abby YongNo ratings yet
- Lecturer 2 Innate Immune System and Adaptve Immune SystemDocument6 pagesLecturer 2 Innate Immune System and Adaptve Immune SystemKevin KipropNo ratings yet
- Immune SystemDocument6 pagesImmune SystemSanNo ratings yet
- The Immune Sytem Phagocytes: Origin & Mode of Action: BloodDocument24 pagesThe Immune Sytem Phagocytes: Origin & Mode of Action: BloodTafadzwaNo ratings yet
- LECTURE 06 Lymphoid System and Location of The Immune CellsDocument16 pagesLECTURE 06 Lymphoid System and Location of The Immune Cellssamarsamar6000No ratings yet
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- Review Flash Card by Dr. Mitch Roy MaristelaDocument205 pagesReview Flash Card by Dr. Mitch Roy MaristelaMark Justin OcampoNo ratings yet
- BIO 425 StudyGuide EXAM IIDocument2 pagesBIO 425 StudyGuide EXAM IIAlexandria BrooksNo ratings yet
- Rajesh Guru Mits OrissaDocument22 pagesRajesh Guru Mits OrissaRajesh GuruNo ratings yet
- Biopsi - M. Fahizza Amru - 111711133110 - 10Document11 pagesBiopsi - M. Fahizza Amru - 111711133110 - 10Fahizza AmruNo ratings yet
- Benign Malignant: Malignant Transformation: Retrovirus (RNA Virus)Document3 pagesBenign Malignant: Malignant Transformation: Retrovirus (RNA Virus)ChiNo ratings yet
- T Cell Maturation, Activation and DifferentiationDocument65 pagesT Cell Maturation, Activation and DifferentiationChanDan RjNo ratings yet
- TransplantationpptfinalDocument57 pagesTransplantationpptfinalmilka berhaneNo ratings yet
- 1 Basic Concepts in ImmunologyDocument31 pages1 Basic Concepts in Immunologyاحمد البزورNo ratings yet
- Immune SystemDocument134 pagesImmune SystemRenz David100% (4)
- Transplantation Immunology: Dr.T.V.Rao MDDocument59 pagesTransplantation Immunology: Dr.T.V.Rao MDtummalapalli venkateswara rao100% (2)
- MRCP NotesDocument33 pagesMRCP NotesHemant DanielNo ratings yet
- Immuno in For MaticsDocument50 pagesImmuno in For MaticsAbraham Omar Espinoza CulupúNo ratings yet
- J Exp Med 1991 Linsley 721 30Document10 pagesJ Exp Med 1991 Linsley 721 30Fernanda RibeiroNo ratings yet
- Major Histocompatibility Complex: Seminar Presented ONDocument19 pagesMajor Histocompatibility Complex: Seminar Presented ONAshikNo ratings yet
- Cell Mediated Immune ResponsesDocument21 pagesCell Mediated Immune ResponsesStefan AlexandruNo ratings yet
- 1b. Antigen & ImunogenDocument13 pages1b. Antigen & ImunogenEl FatihNo ratings yet
- 11 Compressed Notes ImmunityDocument9 pages11 Compressed Notes ImmunityLIM ZHI SHUENNo ratings yet
- Chapter 34 Guyton and HallDocument5 pagesChapter 34 Guyton and Hallg_komolafeNo ratings yet
- Immunology Qs - Part #3, M.TawalbehDocument16 pagesImmunology Qs - Part #3, M.TawalbehMohamed Tawalbe100% (3)
- Score Report - Immunology & Serology at Free Online Quiz SchoolDocument29 pagesScore Report - Immunology & Serology at Free Online Quiz SchoolThomas Giang100% (1)
- Ausarbeitung Immunologie A Korrigierte Fassung WS2019Document185 pagesAusarbeitung Immunologie A Korrigierte Fassung WS2019Adrienn MarkeszNo ratings yet
- 7.2 Immunology Adaptive Immunity 17dec2021Document37 pages7.2 Immunology Adaptive Immunity 17dec20212023406162No ratings yet
- Cluster of Differentiation Antigen PosterDocument1 pageCluster of Differentiation Antigen PosterReeti R. Bhat100% (1)
- Stevens Immuno Sero Answer KeyDocument18 pagesStevens Immuno Sero Answer KeyKATHERINE SHYLE MILARNo ratings yet
- AllergyDocument24 pagesAllergymelNo ratings yet
- Emqs For Medical Students: Second EditionDocument26 pagesEmqs For Medical Students: Second EditionAnonymous ud8gDeN50% (2)
- Test Bank For Human Physiology From Cells To Systems 7th Edition Sherwood DownloadDocument69 pagesTest Bank For Human Physiology From Cells To Systems 7th Edition Sherwood Downloadmichaelwilliamsasrfjgwoym100% (31)
- Schmid 2007Document14 pagesSchmid 2007Camelia AlexandraNo ratings yet