
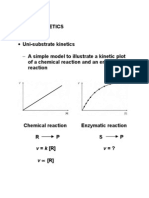
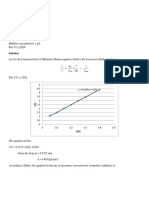
The Michaelis - Menten Equation: Enzyme: Substrate Dissociation Constant. at Equilibrium
The Michaelis - Menten Equation: Enzyme: Substrate Dissociation Constant. at Equilibrium
You might also like
- The Answer Keys Are at The End of The Document.: Section #1 - These Questions Are Worth Two Marks EachDocument26 pagesThe Answer Keys Are at The End of The Document.: Section #1 - These Questions Are Worth Two Marks Eachdsa0% (1)
- IBO Papers (1990-2013)Document3,277 pagesIBO Papers (1990-2013)Science Olympiad Blog90% (10)
- RRKM TheoryDocument4 pagesRRKM TheoryjainankitmumNo ratings yet
- Catálise Enzimática e Energia de GibbsDocument3 pagesCatálise Enzimática e Energia de GibbsRenan OliveiraNo ratings yet
- Calculate KM and Vmax From The Following DataDocument6 pagesCalculate KM and Vmax From The Following DataDr-Dalya Shakir0% (1)
- Lecture 10: Enzyme Kinetics: Reading: Chapter 6 Pp. 202-213Document21 pagesLecture 10: Enzyme Kinetics: Reading: Chapter 6 Pp. 202-213SriArthiNo ratings yet
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- Michaelis Menten EquationDocument4 pagesMichaelis Menten Equationsumant5shringari100% (1)
- Michaelis Menten Equation SummaryDocument7 pagesMichaelis Menten Equation SummaryBüşra GültekinNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsronojoysenguptaNo ratings yet
- Enzyme M&LDocument6 pagesEnzyme M&LNethra Sasikumar100% (1)
- EnzymeKinetics PDFDocument21 pagesEnzymeKinetics PDFaathiraNo ratings yet
- MRQ - #3 Enzyme Kinetics - 2015Document44 pagesMRQ - #3 Enzyme Kinetics - 2015yanuar setiawanNo ratings yet
- Lecture Notes Enzyme 2 Enzyme Kinetics WebDocument29 pagesLecture Notes Enzyme 2 Enzyme Kinetics WebAldren RebaLdeNo ratings yet
- Michaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)Document2 pagesMichaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)fintastellaNo ratings yet
- Enzyme KineticsDocument18 pagesEnzyme KineticsRoyal MindNo ratings yet
- 3 Enzyme KineticsDocument22 pages3 Enzyme KineticsLee Ping Shin100% (1)
- Kjdfks FDocument11 pagesKjdfks FTee BeeNo ratings yet
- Deriving The Michaelis MENTEN EQUTIONDocument5 pagesDeriving The Michaelis MENTEN EQUTIONManish GaurNo ratings yet
- 10-11, Kinetics of Enzyme Catalyzed ReactionDocument34 pages10-11, Kinetics of Enzyme Catalyzed ReactionS. Ansary100% (1)
- Enzymology (GEB-2101) Lecture 3Document15 pagesEnzymology (GEB-2101) Lecture 3Niloy GhoshNo ratings yet
- Question CH06+answer PDFDocument8 pagesQuestion CH06+answer PDFCris-Anne Juangco III100% (1)
- MIT Lecture 6 5.07 Biochemistry LectureDocument16 pagesMIT Lecture 6 5.07 Biochemistry LectureakiridoNo ratings yet
- Enzyme KineticsDocument10 pagesEnzyme KineticsQuenneBelocuraNo ratings yet
- EnzymeDocument39 pagesEnzymeBhaski MadhappanNo ratings yet
- Presentation On BiochemDocument12 pagesPresentation On BiochemRabia HussainNo ratings yet
- Lecture 12 - Enzyme Kinetics IDocument30 pagesLecture 12 - Enzyme Kinetics IThomas Jones50% (2)
- Michaelis-Menten KineticsDocument6 pagesMichaelis-Menten Kineticscharanmann9165No ratings yet
- 2.1 Michaelis Menten EquationDocument8 pages2.1 Michaelis Menten EquationKishoreNo ratings yet
- Kinetics of EnzymesDocument2 pagesKinetics of EnzymesJohnNo ratings yet
- Lab 7 Enzyme KineticsDocument6 pagesLab 7 Enzyme KineticsErikRodriguezNo ratings yet
- Teknik ReaksiDocument21 pagesTeknik ReaksiAhmadi FachryNo ratings yet
- Simple Kinetics of Enzyme ActionDocument7 pagesSimple Kinetics of Enzyme ActionRavindra Mani TiwariNo ratings yet
- Lecture 14: Enzyme Kinetics: Biochemistry I Fall Term, 2004Document6 pagesLecture 14: Enzyme Kinetics: Biochemistry I Fall Term, 2004Louis FortunatoNo ratings yet
- 1 (K) (200 10 10 (3 10 10 (2.3 0.9) Ev (1.60 10) J / Ev 6.64 10Document5 pages1 (K) (200 10 10 (3 10 10 (2.3 0.9) Ev (1.60 10) J / Ev 6.64 10Yant HArdy WaeNo ratings yet
- MichaelisDocument4 pagesMichaelishamzaasghar233No ratings yet
- H C K M) (258 M) M / S) (58 M) J.SDocument5 pagesH C K M) (258 M) M / S) (58 M) J.SYant HArdy WaeNo ratings yet
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Document16 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Ken M'voulaboloNo ratings yet
- Chapter 7.7 Abbreviated Lecture NotesDocument27 pagesChapter 7.7 Abbreviated Lecture NotesZach MaxwellNo ratings yet
- Lecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesDocument21 pagesLecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesAkash AroraNo ratings yet
- Steady-State Microelectrode Voltammetry As A Route To Homogeneous KineticsDocument14 pagesSteady-State Microelectrode Voltammetry As A Route To Homogeneous Kineticsdt5632No ratings yet
- W (U (U (C C HC e DW (:, T), T) - D / D - ) 8 1 1, T) /D 0Document6 pagesW (U (U (C C HC e DW (:, T), T) - D / D - ) 8 1 1, T) /D 0Yant HArdy WaeNo ratings yet
- Electrodinamica CuanticaDocument27 pagesElectrodinamica CuanticaJuan LombarderoNo ratings yet
- Electron Dynamics: E GradDocument6 pagesElectron Dynamics: E GradRashi VermaNo ratings yet
- Michaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiDocument15 pagesMichaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiEmmanuel Alexander PilapilNo ratings yet
- Michaelis Menten DerivedDocument4 pagesMichaelis Menten DerivedGOOGLI TECNo ratings yet
- Salavati Lecture SolutionsDocument3 pagesSalavati Lecture Solutionsanon_900161280No ratings yet
- Classical HamiltonianDocument4 pagesClassical Hamiltonianbatenas1No ratings yet
- LSM1101 Enzyme2Document40 pagesLSM1101 Enzyme2givena2ndchanceNo ratings yet
- Phy Pharm AssignmentDocument4 pagesPhy Pharm AssignmentNazura AliasNo ratings yet
- Griffiths 5.15 16 17 18Document4 pagesGriffiths 5.15 16 17 18Tom smithNo ratings yet
- Michaelis MentenKineticsandBriggs HaldaneKinetics - HTMLDocument2 pagesMichaelis MentenKineticsandBriggs HaldaneKinetics - HTMLCrestore Lex Tapia CapiñaNo ratings yet
- Biophysj00128 0003Document8 pagesBiophysj00128 0003Dean AstumianNo ratings yet
- Enzyme and Acid - Base CatalysisDocument64 pagesEnzyme and Acid - Base Catalysisbinseung skzNo ratings yet
- Bio Chapter 3Document103 pagesBio Chapter 3Areej AlissaNo ratings yet
- Volumetric Perpetual Harmonic OscillatorDocument44 pagesVolumetric Perpetual Harmonic OscillatorDmitry TarasovNo ratings yet
- Lab#06 MSBME11Document6 pagesLab#06 MSBME11BUSHRA KALEEMNo ratings yet
- Chapter 1 Single-Enzyme Kinetics: Molecular Cell BiologyDocument7 pagesChapter 1 Single-Enzyme Kinetics: Molecular Cell BiologySparrowGospleGilbertNo ratings yet
- On The Numerical Computation of Enzyme Kinetic Parameters: Biomath CommunicationsDocument24 pagesOn The Numerical Computation of Enzyme Kinetic Parameters: Biomath CommunicationsNatalia OchoaNo ratings yet
- Biology 3601 Biochemistry Enzyme Kinetics Laboratory BackgroundDocument5 pagesBiology 3601 Biochemistry Enzyme Kinetics Laboratory Backgroundkgeorges27No ratings yet
- EnzomologyDocument26 pagesEnzomologyToga Brandon100% (1)
- ArticleDocument8 pagesArticleSindhu JangamNo ratings yet
- Types of Enzyme Inhibitors Modified Ocober 2022Document45 pagesTypes of Enzyme Inhibitors Modified Ocober 2022DENNIS murageNo ratings yet
- Biocatalysis and Agricultural Biotechnology: Licheniformis ALW1Document9 pagesBiocatalysis and Agricultural Biotechnology: Licheniformis ALW1Nemo NemoNo ratings yet
- Assignment CH62Document3 pagesAssignment CH62Swathi SNo ratings yet
- Enzymes Lecture 1 2015animationDocument80 pagesEnzymes Lecture 1 2015animationSarfaras KKNo ratings yet
- Exp 3 Biochemistry Enzyme ActivityDocument21 pagesExp 3 Biochemistry Enzyme ActivityhasiqmalNo ratings yet
- BIOC 307 Old Exam 1Document17 pagesBIOC 307 Old Exam 1Katie RoseNo ratings yet
- Electrochemical-Enzymatic Determination of Glucose in Beverages - Pine Research Instrumentation StoreDocument20 pagesElectrochemical-Enzymatic Determination of Glucose in Beverages - Pine Research Instrumentation Storeteam TSOTARENo ratings yet
- Lecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesDocument21 pagesLecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesAkash AroraNo ratings yet
- FBE Merged - MidsemDocument276 pagesFBE Merged - MidsemAnusha DesaiNo ratings yet
- Michaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiDocument15 pagesMichaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiEmmanuel Alexander PilapilNo ratings yet
- HOW TO - Calculate KM and Vmax With ExcelDocument3 pagesHOW TO - Calculate KM and Vmax With Excelminjeshp67% (3)
- Nonlinear Pharmacokinetics 1Document11 pagesNonlinear Pharmacokinetics 1Mubashar ShahidNo ratings yet
- PHAR 7633 Chapter 2 Background Mathematical Material: Student Objectives For This ChapterDocument28 pagesPHAR 7633 Chapter 2 Background Mathematical Material: Student Objectives For This ChapterkbnarkhedeNo ratings yet
- Palaganas Hw2 Che 197Document5 pagesPalaganas Hw2 Che 197Elah Palaganas100% (1)
- 3 - Nurizqie Aprilia Iswati - 1811012220018Document10 pages3 - Nurizqie Aprilia Iswati - 1811012220018Nia inayahNo ratings yet
- ESC3508 2 Transformations Into Linear FunctionsDocument7 pagesESC3508 2 Transformations Into Linear FunctionsNUR ATIQAH BINTI MOHD ROSLI / UPMNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsKaren Ivy Bacsain100% (1)
- Pectinase GraphsDocument7 pagesPectinase GraphsanjanakaveriNo ratings yet
- Chemistry Higher Level Paper 3: Instructions To CandidatesDocument40 pagesChemistry Higher Level Paper 3: Instructions To CandidatesAleksander Ziolkowski100% (1)
- Problem Set 2Document5 pagesProblem Set 2Fran LeeNo ratings yet
- Chapter 23Document53 pagesChapter 23haririNo ratings yet
- Enzymes 12 5 19Document113 pagesEnzymes 12 5 19Arshad Mehmood MinhasNo ratings yet
- Exfin Sol PDFDocument11 pagesExfin Sol PDFDavid ValeroNo ratings yet
- Article Lipase in Oil IndustryDocument10 pagesArticle Lipase in Oil Industryyogender aNo ratings yet
- Jurnal Biokimia 1Document10 pagesJurnal Biokimia 1selviNo ratings yet
- Chapter 3:enzymes: Enzymes Lower The Activation Energy of A Reaction byDocument14 pagesChapter 3:enzymes: Enzymes Lower The Activation Energy of A Reaction bySweta GoodurNo ratings yet






















































































Download as doc, pdf, or txt
You might also like
- The Answer Keys Are at The End of The Document.: Section #1 - These Questions Are Worth Two Marks EachDocument26 pagesThe Answer Keys Are at The End of The Document.: Section #1 - These Questions Are Worth Two Marks Eachdsa0% (1)
- IBO Papers (1990-2013)Document3,277 pagesIBO Papers (1990-2013)Science Olympiad Blog90% (10)
- RRKM TheoryDocument4 pagesRRKM TheoryjainankitmumNo ratings yet
- Catálise Enzimática e Energia de GibbsDocument3 pagesCatálise Enzimática e Energia de GibbsRenan OliveiraNo ratings yet
- Calculate KM and Vmax From The Following DataDocument6 pagesCalculate KM and Vmax From The Following DataDr-Dalya Shakir0% (1)
- Lecture 10: Enzyme Kinetics: Reading: Chapter 6 Pp. 202-213Document21 pagesLecture 10: Enzyme Kinetics: Reading: Chapter 6 Pp. 202-213SriArthiNo ratings yet
- EnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Document21 pagesEnzymeKinetics by P.C. Misra Professor, Department of Biochemistry Lucknow University, Lucknow-226 007Dr. SHIVA AITHALNo ratings yet
- Michaelis Menten EquationDocument4 pagesMichaelis Menten Equationsumant5shringari100% (1)
- Michaelis Menten Equation SummaryDocument7 pagesMichaelis Menten Equation SummaryBüşra GültekinNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsronojoysenguptaNo ratings yet
- Enzyme M&LDocument6 pagesEnzyme M&LNethra Sasikumar100% (1)
- EnzymeKinetics PDFDocument21 pagesEnzymeKinetics PDFaathiraNo ratings yet
- MRQ - #3 Enzyme Kinetics - 2015Document44 pagesMRQ - #3 Enzyme Kinetics - 2015yanuar setiawanNo ratings yet
- Lecture Notes Enzyme 2 Enzyme Kinetics WebDocument29 pagesLecture Notes Enzyme 2 Enzyme Kinetics WebAldren RebaLdeNo ratings yet
- Michaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)Document2 pagesMichaelis-Menten Type Kinetics: E S) ES) K K K E S) ES)fintastellaNo ratings yet
- Enzyme KineticsDocument18 pagesEnzyme KineticsRoyal MindNo ratings yet
- 3 Enzyme KineticsDocument22 pages3 Enzyme KineticsLee Ping Shin100% (1)
- Kjdfks FDocument11 pagesKjdfks FTee BeeNo ratings yet
- Deriving The Michaelis MENTEN EQUTIONDocument5 pagesDeriving The Michaelis MENTEN EQUTIONManish GaurNo ratings yet
- 10-11, Kinetics of Enzyme Catalyzed ReactionDocument34 pages10-11, Kinetics of Enzyme Catalyzed ReactionS. Ansary100% (1)
- Enzymology (GEB-2101) Lecture 3Document15 pagesEnzymology (GEB-2101) Lecture 3Niloy GhoshNo ratings yet
- Question CH06+answer PDFDocument8 pagesQuestion CH06+answer PDFCris-Anne Juangco III100% (1)
- MIT Lecture 6 5.07 Biochemistry LectureDocument16 pagesMIT Lecture 6 5.07 Biochemistry LectureakiridoNo ratings yet
- Enzyme KineticsDocument10 pagesEnzyme KineticsQuenneBelocuraNo ratings yet
- EnzymeDocument39 pagesEnzymeBhaski MadhappanNo ratings yet
- Presentation On BiochemDocument12 pagesPresentation On BiochemRabia HussainNo ratings yet
- Lecture 12 - Enzyme Kinetics IDocument30 pagesLecture 12 - Enzyme Kinetics IThomas Jones50% (2)
- Michaelis-Menten KineticsDocument6 pagesMichaelis-Menten Kineticscharanmann9165No ratings yet
- 2.1 Michaelis Menten EquationDocument8 pages2.1 Michaelis Menten EquationKishoreNo ratings yet
- Kinetics of EnzymesDocument2 pagesKinetics of EnzymesJohnNo ratings yet
- Lab 7 Enzyme KineticsDocument6 pagesLab 7 Enzyme KineticsErikRodriguezNo ratings yet
- Teknik ReaksiDocument21 pagesTeknik ReaksiAhmadi FachryNo ratings yet
- Simple Kinetics of Enzyme ActionDocument7 pagesSimple Kinetics of Enzyme ActionRavindra Mani TiwariNo ratings yet
- Lecture 14: Enzyme Kinetics: Biochemistry I Fall Term, 2004Document6 pagesLecture 14: Enzyme Kinetics: Biochemistry I Fall Term, 2004Louis FortunatoNo ratings yet
- 1 (K) (200 10 10 (3 10 10 (2.3 0.9) Ev (1.60 10) J / Ev 6.64 10Document5 pages1 (K) (200 10 10 (3 10 10 (2.3 0.9) Ev (1.60 10) J / Ev 6.64 10Yant HArdy WaeNo ratings yet
- MichaelisDocument4 pagesMichaelishamzaasghar233No ratings yet
- H C K M) (258 M) M / S) (58 M) J.SDocument5 pagesH C K M) (258 M) M / S) (58 M) J.SYant HArdy WaeNo ratings yet
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Document16 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-231-246Ken M'voulaboloNo ratings yet
- Chapter 7.7 Abbreviated Lecture NotesDocument27 pagesChapter 7.7 Abbreviated Lecture NotesZach MaxwellNo ratings yet
- Lecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesDocument21 pagesLecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesAkash AroraNo ratings yet
- Steady-State Microelectrode Voltammetry As A Route To Homogeneous KineticsDocument14 pagesSteady-State Microelectrode Voltammetry As A Route To Homogeneous Kineticsdt5632No ratings yet
- W (U (U (C C HC e DW (:, T), T) - D / D - ) 8 1 1, T) /D 0Document6 pagesW (U (U (C C HC e DW (:, T), T) - D / D - ) 8 1 1, T) /D 0Yant HArdy WaeNo ratings yet
- Electrodinamica CuanticaDocument27 pagesElectrodinamica CuanticaJuan LombarderoNo ratings yet
- Electron Dynamics: E GradDocument6 pagesElectron Dynamics: E GradRashi VermaNo ratings yet
- Michaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiDocument15 pagesMichaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiEmmanuel Alexander PilapilNo ratings yet
- Michaelis Menten DerivedDocument4 pagesMichaelis Menten DerivedGOOGLI TECNo ratings yet
- Salavati Lecture SolutionsDocument3 pagesSalavati Lecture Solutionsanon_900161280No ratings yet
- Classical HamiltonianDocument4 pagesClassical Hamiltonianbatenas1No ratings yet
- LSM1101 Enzyme2Document40 pagesLSM1101 Enzyme2givena2ndchanceNo ratings yet
- Phy Pharm AssignmentDocument4 pagesPhy Pharm AssignmentNazura AliasNo ratings yet
- Griffiths 5.15 16 17 18Document4 pagesGriffiths 5.15 16 17 18Tom smithNo ratings yet
- Michaelis MentenKineticsandBriggs HaldaneKinetics - HTMLDocument2 pagesMichaelis MentenKineticsandBriggs HaldaneKinetics - HTMLCrestore Lex Tapia CapiñaNo ratings yet
- Biophysj00128 0003Document8 pagesBiophysj00128 0003Dean AstumianNo ratings yet
- Enzyme and Acid - Base CatalysisDocument64 pagesEnzyme and Acid - Base Catalysisbinseung skzNo ratings yet
- Bio Chapter 3Document103 pagesBio Chapter 3Areej AlissaNo ratings yet
- Volumetric Perpetual Harmonic OscillatorDocument44 pagesVolumetric Perpetual Harmonic OscillatorDmitry TarasovNo ratings yet
- Lab#06 MSBME11Document6 pagesLab#06 MSBME11BUSHRA KALEEMNo ratings yet
- Chapter 1 Single-Enzyme Kinetics: Molecular Cell BiologyDocument7 pagesChapter 1 Single-Enzyme Kinetics: Molecular Cell BiologySparrowGospleGilbertNo ratings yet
- On The Numerical Computation of Enzyme Kinetic Parameters: Biomath CommunicationsDocument24 pagesOn The Numerical Computation of Enzyme Kinetic Parameters: Biomath CommunicationsNatalia OchoaNo ratings yet
- Biology 3601 Biochemistry Enzyme Kinetics Laboratory BackgroundDocument5 pagesBiology 3601 Biochemistry Enzyme Kinetics Laboratory Backgroundkgeorges27No ratings yet
- EnzomologyDocument26 pagesEnzomologyToga Brandon100% (1)
- ArticleDocument8 pagesArticleSindhu JangamNo ratings yet
- Types of Enzyme Inhibitors Modified Ocober 2022Document45 pagesTypes of Enzyme Inhibitors Modified Ocober 2022DENNIS murageNo ratings yet
- Biocatalysis and Agricultural Biotechnology: Licheniformis ALW1Document9 pagesBiocatalysis and Agricultural Biotechnology: Licheniformis ALW1Nemo NemoNo ratings yet
- Assignment CH62Document3 pagesAssignment CH62Swathi SNo ratings yet
- Enzymes Lecture 1 2015animationDocument80 pagesEnzymes Lecture 1 2015animationSarfaras KKNo ratings yet
- Exp 3 Biochemistry Enzyme ActivityDocument21 pagesExp 3 Biochemistry Enzyme ActivityhasiqmalNo ratings yet
- BIOC 307 Old Exam 1Document17 pagesBIOC 307 Old Exam 1Katie RoseNo ratings yet
- Electrochemical-Enzymatic Determination of Glucose in Beverages - Pine Research Instrumentation StoreDocument20 pagesElectrochemical-Enzymatic Determination of Glucose in Beverages - Pine Research Instrumentation Storeteam TSOTARENo ratings yet
- Lecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesDocument21 pagesLecture Notes On BCH 409: Advanced Enzymology (3 Units) : Enzymes and Life ProcessesAkash AroraNo ratings yet
- FBE Merged - MidsemDocument276 pagesFBE Merged - MidsemAnusha DesaiNo ratings yet
- Michaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiDocument15 pagesMichaelis-Menten Kinetics and Briggs-Haldane Kinetics - ChemwikiEmmanuel Alexander PilapilNo ratings yet
- HOW TO - Calculate KM and Vmax With ExcelDocument3 pagesHOW TO - Calculate KM and Vmax With Excelminjeshp67% (3)
- Nonlinear Pharmacokinetics 1Document11 pagesNonlinear Pharmacokinetics 1Mubashar ShahidNo ratings yet
- PHAR 7633 Chapter 2 Background Mathematical Material: Student Objectives For This ChapterDocument28 pagesPHAR 7633 Chapter 2 Background Mathematical Material: Student Objectives For This ChapterkbnarkhedeNo ratings yet
- Palaganas Hw2 Che 197Document5 pagesPalaganas Hw2 Che 197Elah Palaganas100% (1)
- 3 - Nurizqie Aprilia Iswati - 1811012220018Document10 pages3 - Nurizqie Aprilia Iswati - 1811012220018Nia inayahNo ratings yet
- ESC3508 2 Transformations Into Linear FunctionsDocument7 pagesESC3508 2 Transformations Into Linear FunctionsNUR ATIQAH BINTI MOHD ROSLI / UPMNo ratings yet
- Enzyme KineticsDocument8 pagesEnzyme KineticsKaren Ivy Bacsain100% (1)
- Pectinase GraphsDocument7 pagesPectinase GraphsanjanakaveriNo ratings yet
- Chemistry Higher Level Paper 3: Instructions To CandidatesDocument40 pagesChemistry Higher Level Paper 3: Instructions To CandidatesAleksander Ziolkowski100% (1)
- Problem Set 2Document5 pagesProblem Set 2Fran LeeNo ratings yet
- Chapter 23Document53 pagesChapter 23haririNo ratings yet
- Enzymes 12 5 19Document113 pagesEnzymes 12 5 19Arshad Mehmood MinhasNo ratings yet
- Exfin Sol PDFDocument11 pagesExfin Sol PDFDavid ValeroNo ratings yet
- Article Lipase in Oil IndustryDocument10 pagesArticle Lipase in Oil Industryyogender aNo ratings yet
- Jurnal Biokimia 1Document10 pagesJurnal Biokimia 1selviNo ratings yet
- Chapter 3:enzymes: Enzymes Lower The Activation Energy of A Reaction byDocument14 pagesChapter 3:enzymes: Enzymes Lower The Activation Energy of A Reaction bySweta GoodurNo ratings yet