
Eduardo Blumwald: Sodium Transport and Salt Tolerance in Plants
Eduardo Blumwald: Sodium Transport and Salt Tolerance in Plants
You might also like
- Lesson Plan 5eDocument3 pagesLesson Plan 5eapi-324014008100% (3)
- ASE SC 1 Animal & Plant Cell ColoringDocument3 pagesASE SC 1 Animal & Plant Cell ColoringAlfie Cabario100% (1)
- Plant and Animal CellDocument4 pagesPlant and Animal CellRonald AzoresNo ratings yet
- Cell Revision WorksheetDocument3 pagesCell Revision WorksheetCally Chew100% (5)
- Lab Report Animal and Plant CellsDocument5 pagesLab Report Animal and Plant CellsNicolas Gómez100% (1)
- Intestinal Ion Transport and The Pathophysiology of Diarrhea - PMCDocument22 pagesIntestinal Ion Transport and The Pathophysiology of Diarrhea - PMCtanatswa courtney matingoNo ratings yet
- Differential Diagnosis of Hyperkalemia: An Update To A Complex ProblemDocument9 pagesDifferential Diagnosis of Hyperkalemia: An Update To A Complex ProblemMariana CabralNo ratings yet
- Potassium Transporters in Plants - Involvement in K Acquisition, Redistribution and HomeostasisDocument9 pagesPotassium Transporters in Plants - Involvement in K Acquisition, Redistribution and HomeostasisLuis Felipe RamirezNo ratings yet
- Salt and Water Absorption in The Human Colon: A Modern AppraisalDocument6 pagesSalt and Water Absorption in The Human Colon: A Modern Appraisalurwords2deepNo ratings yet
- WNK ArticuloDocument18 pagesWNK ArticuloangelNo ratings yet
- Mechanism of Salivary GlandDocument5 pagesMechanism of Salivary GlandNirmala M. KarimatannisaNo ratings yet
- Metabolisme KDocument38 pagesMetabolisme KChandra YuniantoNo ratings yet
- Plant Organellar Calcium Signalling: An Emerging FieldDocument18 pagesPlant Organellar Calcium Signalling: An Emerging FieldIndrė LapeikaitėNo ratings yet
- K Channels: Biochemistry of MetabolismDocument23 pagesK Channels: Biochemistry of MetabolismBINDUNo ratings yet
- Intestinal Ion Transport and The Pathophysiology of DiarrheaDocument13 pagesIntestinal Ion Transport and The Pathophysiology of Diarrheajuanfraflores9777No ratings yet
- NM - PCP 2016Document16 pagesNM - PCP 2016aarushNo ratings yet
- Paper No and Title 15: Bioinorganic Chemistry Module No and Title 8: Ion Transport Through Cell Membrane and Na /K Pump Module Tag CHE - P15 - M8Document9 pagesPaper No and Title 15: Bioinorganic Chemistry Module No and Title 8: Ion Transport Through Cell Membrane and Na /K Pump Module Tag CHE - P15 - M8kottooranjohnbNo ratings yet
- 6-Drugs Acting On The Renal System - 2Document65 pages6-Drugs Acting On The Renal System - 2Hamid Hussain HamidNo ratings yet
- KEITH BREWER High PH Therapy For Cancer Tests On Mice and Humans 12PDocument12 pagesKEITH BREWER High PH Therapy For Cancer Tests On Mice and Humans 12PAna LuNo ratings yet
- Assimilasi Sulfur + Gambar: Arabidopsis An Atmrs2/Atmgt Gene Family Encoding MGDocument5 pagesAssimilasi Sulfur + Gambar: Arabidopsis An Atmrs2/Atmgt Gene Family Encoding MGLung KinNo ratings yet
- Cell Signalling Biology - Ion Channels (2012)Document72 pagesCell Signalling Biology - Ion Channels (2012)Apróné Török IbolyaNo ratings yet
- Inorganic and Organic Anion Transport by Insect Renal EpitheliaDocument13 pagesInorganic and Organic Anion Transport by Insect Renal EpitheliaCLPHtheoryNo ratings yet
- NIH Public Access: Author ManuscriptDocument16 pagesNIH Public Access: Author ManuscriptchechenoveliaNo ratings yet
- Calcium Signaling and Cytotoxicity: in MammalianDocument11 pagesCalcium Signaling and Cytotoxicity: in MammalianRoland ToroNo ratings yet
- Ion TransportDocument5 pagesIon TransportSubhaNo ratings yet
- The Plant Journal - 2005 - Ivashikina - K Currents Through SV Type Vacuolar Channels Are Sensitive To Elevated LuminalDocument9 pagesThe Plant Journal - 2005 - Ivashikina - K Currents Through SV Type Vacuolar Channels Are Sensitive To Elevated Luminal38 Sourav BhowmikNo ratings yet
- Ion Channel: 2 Biological RoleDocument8 pagesIon Channel: 2 Biological RoleMartin MayerNo ratings yet
- Ion Channels Are Pore-FormingDocument11 pagesIon Channels Are Pore-FormingshanmukhaNo ratings yet
- Micronutrientes Plantas 2Document11 pagesMicronutrientes Plantas 2Tatiana AndradeNo ratings yet
- Disorders of Potassium BalanceDocument41 pagesDisorders of Potassium BalanceUmesh BabuNo ratings yet
- Nature 03911Document4 pagesNature 03911tit tutNo ratings yet
- 207 Full PDFDocument9 pages207 Full PDFLuis Alberto Alvarado BarzalloNo ratings yet
- Osmo IngDocument11 pagesOsmo IngRara KhentiNo ratings yet
- Lec 14 - Kidney PDFDocument16 pagesLec 14 - Kidney PDFrajeshNo ratings yet
- Renal Tubular Acidosis H Base and Ammonia Transpo 2018 Advances in ChronicDocument17 pagesRenal Tubular Acidosis H Base and Ammonia Transpo 2018 Advances in ChronicSiva RamanNo ratings yet
- ASPB - The Arabidopsis Book PDFDocument1,239 pagesASPB - The Arabidopsis Book PDFGabyMezaCarrascoNo ratings yet
- The Apparent Involvement of Anmes in Mineral Dependent Methane Oxidation, As An Analog For Possible Martian MethanotrophyDocument15 pagesThe Apparent Involvement of Anmes in Mineral Dependent Methane Oxidation, As An Analog For Possible Martian MethanotrophyFurkan Bilgekagan ErtugrulNo ratings yet
- Ferrer2021 Article EnergyCalculationsForPotassiumDocument8 pagesFerrer2021 Article EnergyCalculationsForPotassiumSamuelNo ratings yet
- Extremophilic Microbes Alkaline PHDocument7 pagesExtremophilic Microbes Alkaline PHsashaikh1213No ratings yet
- Salinity Tolerance in PlantsDocument26 pagesSalinity Tolerance in PlantsBrza RajputNo ratings yet
- Pone 0033645Document14 pagesPone 0033645mNo ratings yet
- Content 257-22-13246.fullDocument7 pagesContent 257-22-13246.fullnirmala_amuthaNo ratings yet
- Electrolyte Quintet Potassium PDFDocument6 pagesElectrolyte Quintet Potassium PDFMaria Guadalupe Hernandez Jimenez100% (1)
- Potassium: From Physiology To Clinical Implications: Acid-Base, Electrolyte and Fluid Alterations: ReviewDocument8 pagesPotassium: From Physiology To Clinical Implications: Acid-Base, Electrolyte and Fluid Alterations: Reviewkarlakalua77No ratings yet
- Distributed StructuresDocument10 pagesDistributed StructuresSamuel Morales Navarro100% (1)
- Chloride Channels and Hypercalciuria: An Unturned Stone: Journal of Clinical Investigation April 1997Document4 pagesChloride Channels and Hypercalciuria: An Unturned Stone: Journal of Clinical Investigation April 1997androiide70No ratings yet
- The Electrogenic of SodiumDocument9 pagesThe Electrogenic of SodiumKemuel Ian CometaNo ratings yet
- FISIOLOGI SEL - Raja Pardomuan HarahapDocument60 pagesFISIOLOGI SEL - Raja Pardomuan HarahapRaja PardomuanNo ratings yet
- Cheng HM Key Concepts Priciples Na GradDocument1 pageCheng HM Key Concepts Priciples Na GradAlya LawrentNo ratings yet
- Redução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoDocument5 pagesRedução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoRenan Guilherme de Oliveira GuihNo ratings yet
- Phaseolus Vulgaris, Fully 30 Percent of The Osmotic Potential Change Can Be Accounted For by KDocument3 pagesPhaseolus Vulgaris, Fully 30 Percent of The Osmotic Potential Change Can Be Accounted For by KHamdan FatahNo ratings yet
- Sulfate Radicals Enable A NonDocument13 pagesSulfate Radicals Enable A NonIchon ManadoNo ratings yet
- Calcium Signaling: Encoding and Decoding: Advanced ArticleDocument10 pagesCalcium Signaling: Encoding and Decoding: Advanced ArticleazzaassNo ratings yet
- 1 Phase Transfer ReactionsDocument51 pages1 Phase Transfer ReactionsHiren BhendwalNo ratings yet
- DownloadDocument16 pagesDownloadasalizwa ludlalaNo ratings yet
- 1 Ion Channel - Pertemuan 8-MhsDocument35 pages1 Ion Channel - Pertemuan 8-MhsDwi Amalia husnaNo ratings yet
- T RPV CuatroDocument6 pagesT RPV CuatroJoaquínaLoeraNo ratings yet
- Springer Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Microbial EcologyDocument11 pagesSpringer Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Microbial EcologySherly Kurnia DewiNo ratings yet
- Corona 2016Document10 pagesCorona 2016Vasudev M SNo ratings yet
- Kla DeterminationDocument23 pagesKla DeterminationJokanoe LertNo ratings yet
- Ion TransportersDocument2 pagesIon TransportersfilipemedicinavrNo ratings yet
- Calcium Channels: The Importance of Calcium Channels: Discovery of Voltage-Gated Calcium ChannelsDocument4 pagesCalcium Channels: The Importance of Calcium Channels: Discovery of Voltage-Gated Calcium ChannelsKayleeNo ratings yet
- Organellar Calcium Signaling in PlantsDocument10 pagesOrganellar Calcium Signaling in PlantsasifNo ratings yet
- Membrane Transport: Biochemistry of MetabolismDocument39 pagesMembrane Transport: Biochemistry of MetabolismJonny2010No ratings yet
- Chromans and TocopherolsFrom EverandChromans and TocopherolsGwynn P. EllisNo ratings yet
- B2 2+Organelles+and+Compartmentalization+SL+Document24 pagesB2 2+Organelles+and+Compartmentalization+SL+Jiwoo ParkNo ratings yet
- Ujian 1 Biologi Form 4Document12 pagesUjian 1 Biologi Form 4edain84No ratings yet
- 2.1 - Cell FunctionDocument29 pages2.1 - Cell Functionliefeng81100% (2)
- Cell Structure AND Cell OrganisationDocument54 pagesCell Structure AND Cell OrganisationAngel Cascayan Delos SantosNo ratings yet
- Lysosome S: Dr. Ranjana SethDocument17 pagesLysosome S: Dr. Ranjana SethDid GorgNo ratings yet
- Eukaryotic Cell StrucDocument27 pagesEukaryotic Cell StrucHari. ChadhaNo ratings yet
- Membrane TransportDocument74 pagesMembrane TransportMiftahul KhairaniNo ratings yet
- Juego de InglesDocument4 pagesJuego de InglesPrueba pruebaNo ratings yet
- G10 PB WE CELL STRUCTURE BIO Fixx Bangettt Banget YaaaaDocument7 pagesG10 PB WE CELL STRUCTURE BIO Fixx Bangettt Banget YaaaaLia lylNo ratings yet
- Struktur Sel Dan OrganelDocument33 pagesStruktur Sel Dan OrganelSeptian IksanNo ratings yet
- Interactive Textbook 1 2 PDFDocument7 pagesInteractive Textbook 1 2 PDFapi-240094705No ratings yet
- Refference BiologyDocument41 pagesRefference BiologyRobert Andrew BaldadoNo ratings yet
- Expt in LabDocument4 pagesExpt in LabMark GirasolNo ratings yet
- SPM 2005 & 2006 - Kertas 1 Set 2Document28 pagesSPM 2005 & 2006 - Kertas 1 Set 2kiongocNo ratings yet
- Module I-Cocepts in BiologyDocument46 pagesModule I-Cocepts in BiologyManan MaheshwariNo ratings yet
- Cells and Microscope Practice TestDocument6 pagesCells and Microscope Practice TestlevchapNo ratings yet
- Cell Parts Practice QuizDocument3 pagesCell Parts Practice QuizAnonymous yFS1ZfmBpNo ratings yet
- GCSE Cells Cut and Stick WorksheetDocument5 pagesGCSE Cells Cut and Stick WorksheetSara NahomNo ratings yet
- 9 Practical - 1Document5 pages9 Practical - 1anubh50No ratings yet
- Laboratory Activity 2 Parts of The CellsDocument3 pagesLaboratory Activity 2 Parts of The CellsAndy GacuyaNo ratings yet
- Biology Notes Grade 10-12Document159 pagesBiology Notes Grade 10-12Gabriel Tambwe100% (6)
- Central VacuoleDocument10 pagesCentral VacuoleBlack manNo ratings yet
- FEDNA 2017 NR Pedersen - Actividad Enzimática Por Microscopía PDFDocument33 pagesFEDNA 2017 NR Pedersen - Actividad Enzimática Por Microscopía PDFNinfa PedersenNo ratings yet
- Cornell Notes Unit 5 ActiveDocument2 pagesCornell Notes Unit 5 Activeapi-3300708430% (1)
- Cell AnalogyDocument11 pagesCell AnalogyMikaella Jayne CatanaoanNo ratings yet


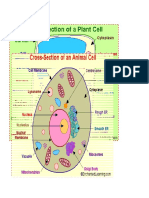






















































































Download as pdf or txt
You might also like
- Lesson Plan 5eDocument3 pagesLesson Plan 5eapi-324014008100% (3)
- ASE SC 1 Animal & Plant Cell ColoringDocument3 pagesASE SC 1 Animal & Plant Cell ColoringAlfie Cabario100% (1)
- Plant and Animal CellDocument4 pagesPlant and Animal CellRonald AzoresNo ratings yet
- Cell Revision WorksheetDocument3 pagesCell Revision WorksheetCally Chew100% (5)
- Lab Report Animal and Plant CellsDocument5 pagesLab Report Animal and Plant CellsNicolas Gómez100% (1)
- Intestinal Ion Transport and The Pathophysiology of Diarrhea - PMCDocument22 pagesIntestinal Ion Transport and The Pathophysiology of Diarrhea - PMCtanatswa courtney matingoNo ratings yet
- Differential Diagnosis of Hyperkalemia: An Update To A Complex ProblemDocument9 pagesDifferential Diagnosis of Hyperkalemia: An Update To A Complex ProblemMariana CabralNo ratings yet
- Potassium Transporters in Plants - Involvement in K Acquisition, Redistribution and HomeostasisDocument9 pagesPotassium Transporters in Plants - Involvement in K Acquisition, Redistribution and HomeostasisLuis Felipe RamirezNo ratings yet
- Salt and Water Absorption in The Human Colon: A Modern AppraisalDocument6 pagesSalt and Water Absorption in The Human Colon: A Modern Appraisalurwords2deepNo ratings yet
- WNK ArticuloDocument18 pagesWNK ArticuloangelNo ratings yet
- Mechanism of Salivary GlandDocument5 pagesMechanism of Salivary GlandNirmala M. KarimatannisaNo ratings yet
- Metabolisme KDocument38 pagesMetabolisme KChandra YuniantoNo ratings yet
- Plant Organellar Calcium Signalling: An Emerging FieldDocument18 pagesPlant Organellar Calcium Signalling: An Emerging FieldIndrė LapeikaitėNo ratings yet
- K Channels: Biochemistry of MetabolismDocument23 pagesK Channels: Biochemistry of MetabolismBINDUNo ratings yet
- Intestinal Ion Transport and The Pathophysiology of DiarrheaDocument13 pagesIntestinal Ion Transport and The Pathophysiology of Diarrheajuanfraflores9777No ratings yet
- NM - PCP 2016Document16 pagesNM - PCP 2016aarushNo ratings yet
- Paper No and Title 15: Bioinorganic Chemistry Module No and Title 8: Ion Transport Through Cell Membrane and Na /K Pump Module Tag CHE - P15 - M8Document9 pagesPaper No and Title 15: Bioinorganic Chemistry Module No and Title 8: Ion Transport Through Cell Membrane and Na /K Pump Module Tag CHE - P15 - M8kottooranjohnbNo ratings yet
- 6-Drugs Acting On The Renal System - 2Document65 pages6-Drugs Acting On The Renal System - 2Hamid Hussain HamidNo ratings yet
- KEITH BREWER High PH Therapy For Cancer Tests On Mice and Humans 12PDocument12 pagesKEITH BREWER High PH Therapy For Cancer Tests On Mice and Humans 12PAna LuNo ratings yet
- Assimilasi Sulfur + Gambar: Arabidopsis An Atmrs2/Atmgt Gene Family Encoding MGDocument5 pagesAssimilasi Sulfur + Gambar: Arabidopsis An Atmrs2/Atmgt Gene Family Encoding MGLung KinNo ratings yet
- Cell Signalling Biology - Ion Channels (2012)Document72 pagesCell Signalling Biology - Ion Channels (2012)Apróné Török IbolyaNo ratings yet
- Inorganic and Organic Anion Transport by Insect Renal EpitheliaDocument13 pagesInorganic and Organic Anion Transport by Insect Renal EpitheliaCLPHtheoryNo ratings yet
- NIH Public Access: Author ManuscriptDocument16 pagesNIH Public Access: Author ManuscriptchechenoveliaNo ratings yet
- Calcium Signaling and Cytotoxicity: in MammalianDocument11 pagesCalcium Signaling and Cytotoxicity: in MammalianRoland ToroNo ratings yet
- Ion TransportDocument5 pagesIon TransportSubhaNo ratings yet
- The Plant Journal - 2005 - Ivashikina - K Currents Through SV Type Vacuolar Channels Are Sensitive To Elevated LuminalDocument9 pagesThe Plant Journal - 2005 - Ivashikina - K Currents Through SV Type Vacuolar Channels Are Sensitive To Elevated Luminal38 Sourav BhowmikNo ratings yet
- Ion Channel: 2 Biological RoleDocument8 pagesIon Channel: 2 Biological RoleMartin MayerNo ratings yet
- Ion Channels Are Pore-FormingDocument11 pagesIon Channels Are Pore-FormingshanmukhaNo ratings yet
- Micronutrientes Plantas 2Document11 pagesMicronutrientes Plantas 2Tatiana AndradeNo ratings yet
- Disorders of Potassium BalanceDocument41 pagesDisorders of Potassium BalanceUmesh BabuNo ratings yet
- Nature 03911Document4 pagesNature 03911tit tutNo ratings yet
- 207 Full PDFDocument9 pages207 Full PDFLuis Alberto Alvarado BarzalloNo ratings yet
- Osmo IngDocument11 pagesOsmo IngRara KhentiNo ratings yet
- Lec 14 - Kidney PDFDocument16 pagesLec 14 - Kidney PDFrajeshNo ratings yet
- Renal Tubular Acidosis H Base and Ammonia Transpo 2018 Advances in ChronicDocument17 pagesRenal Tubular Acidosis H Base and Ammonia Transpo 2018 Advances in ChronicSiva RamanNo ratings yet
- ASPB - The Arabidopsis Book PDFDocument1,239 pagesASPB - The Arabidopsis Book PDFGabyMezaCarrascoNo ratings yet
- The Apparent Involvement of Anmes in Mineral Dependent Methane Oxidation, As An Analog For Possible Martian MethanotrophyDocument15 pagesThe Apparent Involvement of Anmes in Mineral Dependent Methane Oxidation, As An Analog For Possible Martian MethanotrophyFurkan Bilgekagan ErtugrulNo ratings yet
- Ferrer2021 Article EnergyCalculationsForPotassiumDocument8 pagesFerrer2021 Article EnergyCalculationsForPotassiumSamuelNo ratings yet
- Extremophilic Microbes Alkaline PHDocument7 pagesExtremophilic Microbes Alkaline PHsashaikh1213No ratings yet
- Salinity Tolerance in PlantsDocument26 pagesSalinity Tolerance in PlantsBrza RajputNo ratings yet
- Pone 0033645Document14 pagesPone 0033645mNo ratings yet
- Content 257-22-13246.fullDocument7 pagesContent 257-22-13246.fullnirmala_amuthaNo ratings yet
- Electrolyte Quintet Potassium PDFDocument6 pagesElectrolyte Quintet Potassium PDFMaria Guadalupe Hernandez Jimenez100% (1)
- Potassium: From Physiology To Clinical Implications: Acid-Base, Electrolyte and Fluid Alterations: ReviewDocument8 pagesPotassium: From Physiology To Clinical Implications: Acid-Base, Electrolyte and Fluid Alterations: Reviewkarlakalua77No ratings yet
- Distributed StructuresDocument10 pagesDistributed StructuresSamuel Morales Navarro100% (1)
- Chloride Channels and Hypercalciuria: An Unturned Stone: Journal of Clinical Investigation April 1997Document4 pagesChloride Channels and Hypercalciuria: An Unturned Stone: Journal of Clinical Investigation April 1997androiide70No ratings yet
- The Electrogenic of SodiumDocument9 pagesThe Electrogenic of SodiumKemuel Ian CometaNo ratings yet
- FISIOLOGI SEL - Raja Pardomuan HarahapDocument60 pagesFISIOLOGI SEL - Raja Pardomuan HarahapRaja PardomuanNo ratings yet
- Cheng HM Key Concepts Priciples Na GradDocument1 pageCheng HM Key Concepts Priciples Na GradAlya LawrentNo ratings yet
- Redução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoDocument5 pagesRedução de Simetria em Um Canal de Íons Homotetramérico Ativado Por HiperpolarizaçãoRenan Guilherme de Oliveira GuihNo ratings yet
- Phaseolus Vulgaris, Fully 30 Percent of The Osmotic Potential Change Can Be Accounted For by KDocument3 pagesPhaseolus Vulgaris, Fully 30 Percent of The Osmotic Potential Change Can Be Accounted For by KHamdan FatahNo ratings yet
- Sulfate Radicals Enable A NonDocument13 pagesSulfate Radicals Enable A NonIchon ManadoNo ratings yet
- Calcium Signaling: Encoding and Decoding: Advanced ArticleDocument10 pagesCalcium Signaling: Encoding and Decoding: Advanced ArticleazzaassNo ratings yet
- 1 Phase Transfer ReactionsDocument51 pages1 Phase Transfer ReactionsHiren BhendwalNo ratings yet
- DownloadDocument16 pagesDownloadasalizwa ludlalaNo ratings yet
- 1 Ion Channel - Pertemuan 8-MhsDocument35 pages1 Ion Channel - Pertemuan 8-MhsDwi Amalia husnaNo ratings yet
- T RPV CuatroDocument6 pagesT RPV CuatroJoaquínaLoeraNo ratings yet
- Springer Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Microbial EcologyDocument11 pagesSpringer Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Microbial EcologySherly Kurnia DewiNo ratings yet
- Corona 2016Document10 pagesCorona 2016Vasudev M SNo ratings yet
- Kla DeterminationDocument23 pagesKla DeterminationJokanoe LertNo ratings yet
- Ion TransportersDocument2 pagesIon TransportersfilipemedicinavrNo ratings yet
- Calcium Channels: The Importance of Calcium Channels: Discovery of Voltage-Gated Calcium ChannelsDocument4 pagesCalcium Channels: The Importance of Calcium Channels: Discovery of Voltage-Gated Calcium ChannelsKayleeNo ratings yet
- Organellar Calcium Signaling in PlantsDocument10 pagesOrganellar Calcium Signaling in PlantsasifNo ratings yet
- Membrane Transport: Biochemistry of MetabolismDocument39 pagesMembrane Transport: Biochemistry of MetabolismJonny2010No ratings yet
- Chromans and TocopherolsFrom EverandChromans and TocopherolsGwynn P. EllisNo ratings yet
- B2 2+Organelles+and+Compartmentalization+SL+Document24 pagesB2 2+Organelles+and+Compartmentalization+SL+Jiwoo ParkNo ratings yet
- Ujian 1 Biologi Form 4Document12 pagesUjian 1 Biologi Form 4edain84No ratings yet
- 2.1 - Cell FunctionDocument29 pages2.1 - Cell Functionliefeng81100% (2)
- Cell Structure AND Cell OrganisationDocument54 pagesCell Structure AND Cell OrganisationAngel Cascayan Delos SantosNo ratings yet
- Lysosome S: Dr. Ranjana SethDocument17 pagesLysosome S: Dr. Ranjana SethDid GorgNo ratings yet
- Eukaryotic Cell StrucDocument27 pagesEukaryotic Cell StrucHari. ChadhaNo ratings yet
- Membrane TransportDocument74 pagesMembrane TransportMiftahul KhairaniNo ratings yet
- Juego de InglesDocument4 pagesJuego de InglesPrueba pruebaNo ratings yet
- G10 PB WE CELL STRUCTURE BIO Fixx Bangettt Banget YaaaaDocument7 pagesG10 PB WE CELL STRUCTURE BIO Fixx Bangettt Banget YaaaaLia lylNo ratings yet
- Struktur Sel Dan OrganelDocument33 pagesStruktur Sel Dan OrganelSeptian IksanNo ratings yet
- Interactive Textbook 1 2 PDFDocument7 pagesInteractive Textbook 1 2 PDFapi-240094705No ratings yet
- Refference BiologyDocument41 pagesRefference BiologyRobert Andrew BaldadoNo ratings yet
- Expt in LabDocument4 pagesExpt in LabMark GirasolNo ratings yet
- SPM 2005 & 2006 - Kertas 1 Set 2Document28 pagesSPM 2005 & 2006 - Kertas 1 Set 2kiongocNo ratings yet
- Module I-Cocepts in BiologyDocument46 pagesModule I-Cocepts in BiologyManan MaheshwariNo ratings yet
- Cells and Microscope Practice TestDocument6 pagesCells and Microscope Practice TestlevchapNo ratings yet
- Cell Parts Practice QuizDocument3 pagesCell Parts Practice QuizAnonymous yFS1ZfmBpNo ratings yet
- GCSE Cells Cut and Stick WorksheetDocument5 pagesGCSE Cells Cut and Stick WorksheetSara NahomNo ratings yet
- 9 Practical - 1Document5 pages9 Practical - 1anubh50No ratings yet
- Laboratory Activity 2 Parts of The CellsDocument3 pagesLaboratory Activity 2 Parts of The CellsAndy GacuyaNo ratings yet
- Biology Notes Grade 10-12Document159 pagesBiology Notes Grade 10-12Gabriel Tambwe100% (6)
- Central VacuoleDocument10 pagesCentral VacuoleBlack manNo ratings yet
- FEDNA 2017 NR Pedersen - Actividad Enzimática Por Microscopía PDFDocument33 pagesFEDNA 2017 NR Pedersen - Actividad Enzimática Por Microscopía PDFNinfa PedersenNo ratings yet
- Cornell Notes Unit 5 ActiveDocument2 pagesCornell Notes Unit 5 Activeapi-3300708430% (1)
- Cell AnalogyDocument11 pagesCell AnalogyMikaella Jayne CatanaoanNo ratings yet