
Fito Bahan
Fito Bahan
You might also like
- Purine Metabolism de Novo Synthesis and Salvage Pathway, 2015Document28 pagesPurine Metabolism de Novo Synthesis and Salvage Pathway, 2015Tehreem NadeemNo ratings yet
- Mevalonate PathwayDocument13 pagesMevalonate Pathwayfitriani fajriNo ratings yet
- OxidatveDocument10 pagesOxidatvehurainsahar21No ratings yet
- Purine Metabolism PDFDocument29 pagesPurine Metabolism PDFtrinitysugumar0% (1)
- Sodium K PumpDocument11 pagesSodium K Pumphurainsahar21No ratings yet
- CurrPharmDesign2007 DXR InhibitorsDocument18 pagesCurrPharmDesign2007 DXR InhibitorsVincent GeruszNo ratings yet
- PG Purine and PyrimidineDocument14 pagesPG Purine and PyrimidineAbdulnafiu AhmadNo ratings yet
- Acetate, Mevalonate and Shikimic Acid PathwaysDocument19 pagesAcetate, Mevalonate and Shikimic Acid Pathwaysamitaggarwal7888% (8)
- Biosynthetic Pathways - GPDocument46 pagesBiosynthetic Pathways - GPGhanshyam R ParmarNo ratings yet
- Carotenoid: Carotenoids (/KDocument18 pagesCarotenoid: Carotenoids (/Khasan jamiNo ratings yet
- Oxidative PhosphorylationDocument33 pagesOxidative PhosphorylationJithendra Babu0% (1)
- S Adenosylmethionine PDFDocument7 pagesS Adenosylmethionine PDFmanoj_rkl_07No ratings yet
- Enzymes ReglatonDocument11 pagesEnzymes Reglatonhurainsahar21No ratings yet
- ATP Energy Currency of CellDocument5 pagesATP Energy Currency of CellvachhanikapilNo ratings yet
- Atp CycleDocument4 pagesAtp CycleVignesh100% (1)
- Cellular RespirationDocument27 pagesCellular Respirationhurainsahar21No ratings yet
- Purine Pyrimidine BiosynthesisDocument13 pagesPurine Pyrimidine Biosynthesisshanto.tn98No ratings yet
- Embden Meyerhof Parnas & Gluconeogenesis Pathways PDFDocument23 pagesEmbden Meyerhof Parnas & Gluconeogenesis Pathways PDFزهراء فاضل اجبير فعيلNo ratings yet
- ATP: Universal Currency of Cellular Energy: ATP Structure and FunctionDocument12 pagesATP: Universal Currency of Cellular Energy: ATP Structure and FunctionCielo PepitoNo ratings yet
- Labdanes of Natural Origin-Biological ActivitiesDocument24 pagesLabdanes of Natural Origin-Biological Activitiestheyuri@tlen.plNo ratings yet
- Explain The Complete Reaction For The Hydrolysis of Adenosine Triphosphat1Document15 pagesExplain The Complete Reaction For The Hydrolysis of Adenosine Triphosphat1NURAIHAN BINTI HASHIM MoeNo ratings yet
- Mouse Thymidylate Synthase Does Not Show The Inact 2016Document8 pagesMouse Thymidylate Synthase Does Not Show The Inact 2016adamjarmula20No ratings yet
- Chapter 3bDocument15 pagesChapter 3bVim EnobioNo ratings yet
- Fosfatase Soluvel e LeismaniaDocument8 pagesFosfatase Soluvel e LeismaniacoringacravoNo ratings yet
- MitocondriDocument12 pagesMitocondrialberto_uniNo ratings yet
- Gen Bio Mod 9-11Document7 pagesGen Bio Mod 9-11Arabella BrionesNo ratings yet
- Plasmodium Falciparum: Crosstalk Between Purine Nucleotide Metabolism and Mitochondrial Pathways inDocument10 pagesPlasmodium Falciparum: Crosstalk Between Purine Nucleotide Metabolism and Mitochondrial Pathways inayuNo ratings yet
- Metabolism of NucleotidesDocument24 pagesMetabolism of NucleotidesAditya NayakNo ratings yet
- Precursor-Directed Biosynthesis of Prenylated Polyphenols and CarbazolesDocument3 pagesPrecursor-Directed Biosynthesis of Prenylated Polyphenols and CarbazolesNur ChamidahNo ratings yet
- Microbiology With Diseases by Taxonomy 5th Edition Bauman Solutions Manual 1Document36 pagesMicrobiology With Diseases by Taxonomy 5th Edition Bauman Solutions Manual 1tinabrooksidzrkgytcx100% (29)
- Intermediary MetabolismDocument11 pagesIntermediary Metabolismsoumya palavalasaNo ratings yet
- Mitochondrial Diseases and Genetic Defects of ATP SynthaseDocument6 pagesMitochondrial Diseases and Genetic Defects of ATP SynthasewlymjzdwNo ratings yet
- Atp: Universal Currency of Cellular Energy: - Aditya Sunil Nair - Class XI D - Roll No: 22Document10 pagesAtp: Universal Currency of Cellular Energy: - Aditya Sunil Nair - Class XI D - Roll No: 22Aditya Sunil NairNo ratings yet
- U-5 Energy Transformation PDFDocument41 pagesU-5 Energy Transformation PDFHawi kelbessaNo ratings yet
- ATP Synthesis and Storage: Original ArticleDocument15 pagesATP Synthesis and Storage: Original ArticleRiley RilanNo ratings yet
- Bacterial Fermentation NatureDocument7 pagesBacterial Fermentation NatureBayu PutraNo ratings yet
- Gen Bio Cycles - FermentationDocument13 pagesGen Bio Cycles - Fermentationyxcz.rzNo ratings yet
- s41467 019 13766 2 PDFDocument11 pagess41467 019 13766 2 PDFCarolina RicárdezNo ratings yet
- Atp SynthesisDocument5 pagesAtp Synthesiskman0722No ratings yet
- 05-Terpenos y EsteroidesDocument23 pages05-Terpenos y EsteroidesAle VillaNo ratings yet
- Atp Adp CycleDocument13 pagesAtp Adp Cycleelladomingo54No ratings yet
- Research Project: Prof. Dr. Rehab HamedDocument13 pagesResearch Project: Prof. Dr. Rehab HamedHassaninElaradyNo ratings yet
- NOTES Tutorial 4 Proteins RevisionNotes BMSC2120Document5 pagesNOTES Tutorial 4 Proteins RevisionNotes BMSC2120ZAFFAR QURESHINo ratings yet
- BCH 313 Lecture NoteDocument15 pagesBCH 313 Lecture NoteaualbidawyNo ratings yet
- Unit Five Last One Grade 11 Biology Energy TransformationDocument55 pagesUnit Five Last One Grade 11 Biology Energy TransformationaxumfunNo ratings yet
- Steps of GlycolysisDocument3 pagesSteps of GlycolysisIm jungkook JUSTIN SEAGULL A.K.A jungshookNo ratings yet
- Thiocyanate Modulates The Catalytic Activity of Mammalian PeroxidasesDocument9 pagesThiocyanate Modulates The Catalytic Activity of Mammalian PeroxidasesRudolf KiraljNo ratings yet
- ATp HyrolysisDocument9 pagesATp Hyrolysishurainsahar21No ratings yet
- Lec32 F08 HandoutDocument11 pagesLec32 F08 HandoutSarayu Ragavan NairNo ratings yet
- Bio Chap 10Document6 pagesBio Chap 10Nethra SasikumarNo ratings yet
- GLYCOLYSIS A TEN STEP PROCESSDocument3 pagesGLYCOLYSIS A TEN STEP PROCESSSophia ManzanoNo ratings yet
- Lecture 3Document100 pagesLecture 36002 AJAY KUMAR KNo ratings yet
- Glycolysis: A Narrative Summary: Calabano, Mark Jasper C. Biochemistry Lecture II-BS Medical TechnologyDocument2 pagesGlycolysis: A Narrative Summary: Calabano, Mark Jasper C. Biochemistry Lecture II-BS Medical TechnologyMarkJasperCalabanoNo ratings yet
- Q2 Reviewer in General BiologyDocument7 pagesQ2 Reviewer in General BiologyariannealzagaNo ratings yet
- ADENOSINE TRIPHOSPHATE-editDocument30 pagesADENOSINE TRIPHOSPHATE-editSiti Ayu MegawatiNo ratings yet
- Glycolysis Part 3Document12 pagesGlycolysis Part 3Rishav BajoriaNo ratings yet
- STUDY GUIDE 1 NotesDocument4 pagesSTUDY GUIDE 1 NotesBae SeujiNo ratings yet
- Terpenos BiosintesisDocument12 pagesTerpenos BiosintesisOctavio CarbajalNo ratings yet
- Þ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCDocument12 pagesÞ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCgauravNo ratings yet
- Matrix Metalloproteinase BiologyFrom EverandMatrix Metalloproteinase BiologyIrit SagiNo ratings yet
- VICTREX CT 100 Polymer - Technical Data SheetDocument2 pagesVICTREX CT 100 Polymer - Technical Data SheetHari KiranNo ratings yet
- Reaffirmed 1996Document9 pagesReaffirmed 1996Vineet VermaNo ratings yet
- 21Document32 pages21ziadddNo ratings yet
- Evaporator SelectionDocument4 pagesEvaporator SelectionHAIMANOTNo ratings yet
- MSC Semester Iv 16P4Chet14El Advanced Organic Chemistry: Dr. V.S SebastianDocument42 pagesMSC Semester Iv 16P4Chet14El Advanced Organic Chemistry: Dr. V.S SebastianDhruv JariwalaNo ratings yet
- Ads - 2019 - StarTab - Content Uniformity - FinalDocument3 pagesAds - 2019 - StarTab - Content Uniformity - FinalCesar Hernández RamírezNo ratings yet
- Lecture 6 Riviting DesignDocument17 pagesLecture 6 Riviting DesignAbdelrahman yasser ZaghloulNo ratings yet
- AcidsBases and PHDocument6 pagesAcidsBases and PHroslinazriNo ratings yet
- SdarticleDocument9 pagesSdarticleSherif James KaldasNo ratings yet
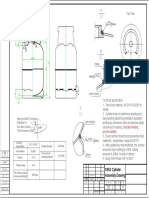
- 10KG Cylinder Assembly DrawingDocument1 page10KG Cylinder Assembly Drawingpaul akhanobaNo ratings yet
- Determination of FatDocument5 pagesDetermination of Fatkolita kamal100% (4)
- TPD ABCIL CAUSTIC PLANT MANUAL RevDocument42 pagesTPD ABCIL CAUSTIC PLANT MANUAL Revjavier pividoriNo ratings yet
- IMO55 2021 T2 Problems EngDocument15 pagesIMO55 2021 T2 Problems EngTrần Phạm Gia BảoNo ratings yet
- An Assignment On Human Physiology.Document7 pagesAn Assignment On Human Physiology.Samira RahmanNo ratings yet
- Ahmes Secondary School: Form Six Weekly Examinations Aprily 2020 132/2 Chemistry 2Document5 pagesAhmes Secondary School: Form Six Weekly Examinations Aprily 2020 132/2 Chemistry 2LastschoolkingZNo ratings yet
- Museum ConservationDocument4 pagesMuseum ConservationRitikNo ratings yet
- Week 5 Symbols Letters and ApplicationDocument55 pagesWeek 5 Symbols Letters and ApplicationOnii chan RhejNo ratings yet
- Organic Name Reactions: Nutshell Review & Preview ofDocument9 pagesOrganic Name Reactions: Nutshell Review & Preview ofSai YashwanthNo ratings yet
- Cell Disruption 2Document14 pagesCell Disruption 2Sukanthan RNo ratings yet
- Recommended Oil Cleanliness Level: Application Oil Cleanliness Required in Accordance With ISO 4406Document2 pagesRecommended Oil Cleanliness Level: Application Oil Cleanliness Required in Accordance With ISO 4406sssNo ratings yet
- Chapter 5 Lithography - IDocument22 pagesChapter 5 Lithography - Isatyam singhalNo ratings yet
- Formulation and Characterization of Solid Dispersion of Nisoldipine by Solvent Evaporation MethodDocument8 pagesFormulation and Characterization of Solid Dispersion of Nisoldipine by Solvent Evaporation Methoddini hanifaNo ratings yet
- List of ReferencesDocument17 pagesList of ReferencesYash SankrityayanNo ratings yet
- CP 10 - Factors Affectinging PhotosynthesisDocument3 pagesCP 10 - Factors Affectinging Photosynthesisrifu91No ratings yet
- 2 Phase Vertical Separator Sizing Calculation-AUHDocument26 pages2 Phase Vertical Separator Sizing Calculation-AUHMike Dukas100% (1)
- 03 Bleach Oxidation HandoutDocument6 pages03 Bleach Oxidation HandoutBrian PermejoNo ratings yet
- 8.334: StatMech II Problem Set #2Document2 pages8.334: StatMech II Problem Set #2bidsitlovNo ratings yet
- Naming Inorganic CompoundsDocument18 pagesNaming Inorganic Compoundsaika smithNo ratings yet
- Mole RatioDocument12 pagesMole Ratiomaysanati2007No ratings yet
























































































Download as docx, pdf, or txt
You might also like
- Purine Metabolism de Novo Synthesis and Salvage Pathway, 2015Document28 pagesPurine Metabolism de Novo Synthesis and Salvage Pathway, 2015Tehreem NadeemNo ratings yet
- Mevalonate PathwayDocument13 pagesMevalonate Pathwayfitriani fajriNo ratings yet
- OxidatveDocument10 pagesOxidatvehurainsahar21No ratings yet
- Purine Metabolism PDFDocument29 pagesPurine Metabolism PDFtrinitysugumar0% (1)
- Sodium K PumpDocument11 pagesSodium K Pumphurainsahar21No ratings yet
- CurrPharmDesign2007 DXR InhibitorsDocument18 pagesCurrPharmDesign2007 DXR InhibitorsVincent GeruszNo ratings yet
- PG Purine and PyrimidineDocument14 pagesPG Purine and PyrimidineAbdulnafiu AhmadNo ratings yet
- Acetate, Mevalonate and Shikimic Acid PathwaysDocument19 pagesAcetate, Mevalonate and Shikimic Acid Pathwaysamitaggarwal7888% (8)
- Biosynthetic Pathways - GPDocument46 pagesBiosynthetic Pathways - GPGhanshyam R ParmarNo ratings yet
- Carotenoid: Carotenoids (/KDocument18 pagesCarotenoid: Carotenoids (/Khasan jamiNo ratings yet
- Oxidative PhosphorylationDocument33 pagesOxidative PhosphorylationJithendra Babu0% (1)
- S Adenosylmethionine PDFDocument7 pagesS Adenosylmethionine PDFmanoj_rkl_07No ratings yet
- Enzymes ReglatonDocument11 pagesEnzymes Reglatonhurainsahar21No ratings yet
- ATP Energy Currency of CellDocument5 pagesATP Energy Currency of CellvachhanikapilNo ratings yet
- Atp CycleDocument4 pagesAtp CycleVignesh100% (1)
- Cellular RespirationDocument27 pagesCellular Respirationhurainsahar21No ratings yet
- Purine Pyrimidine BiosynthesisDocument13 pagesPurine Pyrimidine Biosynthesisshanto.tn98No ratings yet
- Embden Meyerhof Parnas & Gluconeogenesis Pathways PDFDocument23 pagesEmbden Meyerhof Parnas & Gluconeogenesis Pathways PDFزهراء فاضل اجبير فعيلNo ratings yet
- ATP: Universal Currency of Cellular Energy: ATP Structure and FunctionDocument12 pagesATP: Universal Currency of Cellular Energy: ATP Structure and FunctionCielo PepitoNo ratings yet
- Labdanes of Natural Origin-Biological ActivitiesDocument24 pagesLabdanes of Natural Origin-Biological Activitiestheyuri@tlen.plNo ratings yet
- Explain The Complete Reaction For The Hydrolysis of Adenosine Triphosphat1Document15 pagesExplain The Complete Reaction For The Hydrolysis of Adenosine Triphosphat1NURAIHAN BINTI HASHIM MoeNo ratings yet
- Mouse Thymidylate Synthase Does Not Show The Inact 2016Document8 pagesMouse Thymidylate Synthase Does Not Show The Inact 2016adamjarmula20No ratings yet
- Chapter 3bDocument15 pagesChapter 3bVim EnobioNo ratings yet
- Fosfatase Soluvel e LeismaniaDocument8 pagesFosfatase Soluvel e LeismaniacoringacravoNo ratings yet
- MitocondriDocument12 pagesMitocondrialberto_uniNo ratings yet
- Gen Bio Mod 9-11Document7 pagesGen Bio Mod 9-11Arabella BrionesNo ratings yet
- Plasmodium Falciparum: Crosstalk Between Purine Nucleotide Metabolism and Mitochondrial Pathways inDocument10 pagesPlasmodium Falciparum: Crosstalk Between Purine Nucleotide Metabolism and Mitochondrial Pathways inayuNo ratings yet
- Metabolism of NucleotidesDocument24 pagesMetabolism of NucleotidesAditya NayakNo ratings yet
- Precursor-Directed Biosynthesis of Prenylated Polyphenols and CarbazolesDocument3 pagesPrecursor-Directed Biosynthesis of Prenylated Polyphenols and CarbazolesNur ChamidahNo ratings yet
- Microbiology With Diseases by Taxonomy 5th Edition Bauman Solutions Manual 1Document36 pagesMicrobiology With Diseases by Taxonomy 5th Edition Bauman Solutions Manual 1tinabrooksidzrkgytcx100% (29)
- Intermediary MetabolismDocument11 pagesIntermediary Metabolismsoumya palavalasaNo ratings yet
- Mitochondrial Diseases and Genetic Defects of ATP SynthaseDocument6 pagesMitochondrial Diseases and Genetic Defects of ATP SynthasewlymjzdwNo ratings yet
- Atp: Universal Currency of Cellular Energy: - Aditya Sunil Nair - Class XI D - Roll No: 22Document10 pagesAtp: Universal Currency of Cellular Energy: - Aditya Sunil Nair - Class XI D - Roll No: 22Aditya Sunil NairNo ratings yet
- U-5 Energy Transformation PDFDocument41 pagesU-5 Energy Transformation PDFHawi kelbessaNo ratings yet
- ATP Synthesis and Storage: Original ArticleDocument15 pagesATP Synthesis and Storage: Original ArticleRiley RilanNo ratings yet
- Bacterial Fermentation NatureDocument7 pagesBacterial Fermentation NatureBayu PutraNo ratings yet
- Gen Bio Cycles - FermentationDocument13 pagesGen Bio Cycles - Fermentationyxcz.rzNo ratings yet
- s41467 019 13766 2 PDFDocument11 pagess41467 019 13766 2 PDFCarolina RicárdezNo ratings yet
- Atp SynthesisDocument5 pagesAtp Synthesiskman0722No ratings yet
- 05-Terpenos y EsteroidesDocument23 pages05-Terpenos y EsteroidesAle VillaNo ratings yet
- Atp Adp CycleDocument13 pagesAtp Adp Cycleelladomingo54No ratings yet
- Research Project: Prof. Dr. Rehab HamedDocument13 pagesResearch Project: Prof. Dr. Rehab HamedHassaninElaradyNo ratings yet
- NOTES Tutorial 4 Proteins RevisionNotes BMSC2120Document5 pagesNOTES Tutorial 4 Proteins RevisionNotes BMSC2120ZAFFAR QURESHINo ratings yet
- BCH 313 Lecture NoteDocument15 pagesBCH 313 Lecture NoteaualbidawyNo ratings yet
- Unit Five Last One Grade 11 Biology Energy TransformationDocument55 pagesUnit Five Last One Grade 11 Biology Energy TransformationaxumfunNo ratings yet
- Steps of GlycolysisDocument3 pagesSteps of GlycolysisIm jungkook JUSTIN SEAGULL A.K.A jungshookNo ratings yet
- Thiocyanate Modulates The Catalytic Activity of Mammalian PeroxidasesDocument9 pagesThiocyanate Modulates The Catalytic Activity of Mammalian PeroxidasesRudolf KiraljNo ratings yet
- ATp HyrolysisDocument9 pagesATp Hyrolysishurainsahar21No ratings yet
- Lec32 F08 HandoutDocument11 pagesLec32 F08 HandoutSarayu Ragavan NairNo ratings yet
- Bio Chap 10Document6 pagesBio Chap 10Nethra SasikumarNo ratings yet
- GLYCOLYSIS A TEN STEP PROCESSDocument3 pagesGLYCOLYSIS A TEN STEP PROCESSSophia ManzanoNo ratings yet
- Lecture 3Document100 pagesLecture 36002 AJAY KUMAR KNo ratings yet
- Glycolysis: A Narrative Summary: Calabano, Mark Jasper C. Biochemistry Lecture II-BS Medical TechnologyDocument2 pagesGlycolysis: A Narrative Summary: Calabano, Mark Jasper C. Biochemistry Lecture II-BS Medical TechnologyMarkJasperCalabanoNo ratings yet
- Q2 Reviewer in General BiologyDocument7 pagesQ2 Reviewer in General BiologyariannealzagaNo ratings yet
- ADENOSINE TRIPHOSPHATE-editDocument30 pagesADENOSINE TRIPHOSPHATE-editSiti Ayu MegawatiNo ratings yet
- Glycolysis Part 3Document12 pagesGlycolysis Part 3Rishav BajoriaNo ratings yet
- STUDY GUIDE 1 NotesDocument4 pagesSTUDY GUIDE 1 NotesBae SeujiNo ratings yet
- Terpenos BiosintesisDocument12 pagesTerpenos BiosintesisOctavio CarbajalNo ratings yet
- Þ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCDocument12 pagesÞ CC) C' (CC& +C++C: CCCCCCCCCCCCCCCCCCCCCCCCCCCCCCgauravNo ratings yet
- Matrix Metalloproteinase BiologyFrom EverandMatrix Metalloproteinase BiologyIrit SagiNo ratings yet
- VICTREX CT 100 Polymer - Technical Data SheetDocument2 pagesVICTREX CT 100 Polymer - Technical Data SheetHari KiranNo ratings yet
- Reaffirmed 1996Document9 pagesReaffirmed 1996Vineet VermaNo ratings yet
- 21Document32 pages21ziadddNo ratings yet
- Evaporator SelectionDocument4 pagesEvaporator SelectionHAIMANOTNo ratings yet
- MSC Semester Iv 16P4Chet14El Advanced Organic Chemistry: Dr. V.S SebastianDocument42 pagesMSC Semester Iv 16P4Chet14El Advanced Organic Chemistry: Dr. V.S SebastianDhruv JariwalaNo ratings yet
- Ads - 2019 - StarTab - Content Uniformity - FinalDocument3 pagesAds - 2019 - StarTab - Content Uniformity - FinalCesar Hernández RamírezNo ratings yet
- Lecture 6 Riviting DesignDocument17 pagesLecture 6 Riviting DesignAbdelrahman yasser ZaghloulNo ratings yet
- AcidsBases and PHDocument6 pagesAcidsBases and PHroslinazriNo ratings yet
- SdarticleDocument9 pagesSdarticleSherif James KaldasNo ratings yet
- 10KG Cylinder Assembly DrawingDocument1 page10KG Cylinder Assembly Drawingpaul akhanobaNo ratings yet
- Determination of FatDocument5 pagesDetermination of Fatkolita kamal100% (4)
- TPD ABCIL CAUSTIC PLANT MANUAL RevDocument42 pagesTPD ABCIL CAUSTIC PLANT MANUAL Revjavier pividoriNo ratings yet
- IMO55 2021 T2 Problems EngDocument15 pagesIMO55 2021 T2 Problems EngTrần Phạm Gia BảoNo ratings yet
- An Assignment On Human Physiology.Document7 pagesAn Assignment On Human Physiology.Samira RahmanNo ratings yet
- Ahmes Secondary School: Form Six Weekly Examinations Aprily 2020 132/2 Chemistry 2Document5 pagesAhmes Secondary School: Form Six Weekly Examinations Aprily 2020 132/2 Chemistry 2LastschoolkingZNo ratings yet
- Museum ConservationDocument4 pagesMuseum ConservationRitikNo ratings yet
- Week 5 Symbols Letters and ApplicationDocument55 pagesWeek 5 Symbols Letters and ApplicationOnii chan RhejNo ratings yet
- Organic Name Reactions: Nutshell Review & Preview ofDocument9 pagesOrganic Name Reactions: Nutshell Review & Preview ofSai YashwanthNo ratings yet
- Cell Disruption 2Document14 pagesCell Disruption 2Sukanthan RNo ratings yet
- Recommended Oil Cleanliness Level: Application Oil Cleanliness Required in Accordance With ISO 4406Document2 pagesRecommended Oil Cleanliness Level: Application Oil Cleanliness Required in Accordance With ISO 4406sssNo ratings yet
- Chapter 5 Lithography - IDocument22 pagesChapter 5 Lithography - Isatyam singhalNo ratings yet
- Formulation and Characterization of Solid Dispersion of Nisoldipine by Solvent Evaporation MethodDocument8 pagesFormulation and Characterization of Solid Dispersion of Nisoldipine by Solvent Evaporation Methoddini hanifaNo ratings yet
- List of ReferencesDocument17 pagesList of ReferencesYash SankrityayanNo ratings yet
- CP 10 - Factors Affectinging PhotosynthesisDocument3 pagesCP 10 - Factors Affectinging Photosynthesisrifu91No ratings yet
- 2 Phase Vertical Separator Sizing Calculation-AUHDocument26 pages2 Phase Vertical Separator Sizing Calculation-AUHMike Dukas100% (1)
- 03 Bleach Oxidation HandoutDocument6 pages03 Bleach Oxidation HandoutBrian PermejoNo ratings yet
- 8.334: StatMech II Problem Set #2Document2 pages8.334: StatMech II Problem Set #2bidsitlovNo ratings yet
- Naming Inorganic CompoundsDocument18 pagesNaming Inorganic Compoundsaika smithNo ratings yet
- Mole RatioDocument12 pagesMole Ratiomaysanati2007No ratings yet