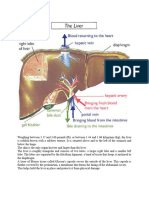
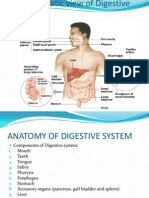
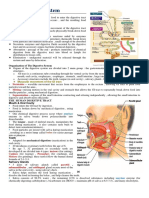
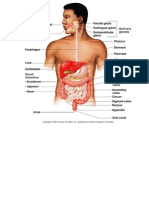
Download as docx, pdf, or txt
You might also like
- The Digestive System A Level NotesDocument7 pagesThe Digestive System A Level Noteselendria100% (1)
- Blood Brain Barrier PDFDocument6 pagesBlood Brain Barrier PDFmanoj_rkl_07100% (1)
- Review PathologyDocument1,175 pagesReview Pathologykandeepan100% (10)
- 6.1 - DigestionDocument8 pages6.1 - DigestionPraveen KhandelwalNo ratings yet
- Human PhysiologyDocument43 pagesHuman PhysiologyBlowDatZaZaNo ratings yet
- Human PhysiologyDocument41 pagesHuman PhysiologyBoris-Jeremie-Latyr GueyeNo ratings yet
- Nutrition and DigestionDocument8 pagesNutrition and DigestionjeffchegzodNo ratings yet
- Digestion Is The Breakdown of Large, Insoluble Food Molecules Into Small, Water-Soluble Molecules Using Mechanical and Chemical ProcessesDocument14 pagesDigestion Is The Breakdown of Large, Insoluble Food Molecules Into Small, Water-Soluble Molecules Using Mechanical and Chemical ProcessesMuhammad AbdullahNo ratings yet
- Further Digestion Absorption Assimilation and Hormones Final QsDocument1 pageFurther Digestion Absorption Assimilation and Hormones Final QsChris WongNo ratings yet
- Human PhysiologyDocument39 pagesHuman PhysiologyML6132021 KENG MING HUI CARMENNo ratings yet
- Structure and Function of The Digestive SystemDocument4 pagesStructure and Function of The Digestive Systemangelignacio2209No ratings yet
- Pencernaan KudaDocument37 pagesPencernaan KudaAchmad FirdaNo ratings yet
- Bio LCV 04 Digestion.3 W17Document45 pagesBio LCV 04 Digestion.3 W17Robert H. GoldsteinNo ratings yet
- SCIENCE 8 Wk. 1Document14 pagesSCIENCE 8 Wk. 1oreooomingNo ratings yet
- DisgestionDocument1 pageDisgestionandressookdeo29No ratings yet
- Name - Digestive System Worksheet: Salivary GlandsDocument5 pagesName - Digestive System Worksheet: Salivary GlandsRJ CamarilloNo ratings yet
- Calamba Doctors' College: Digestion, Absorption, and Metabolism Lesson 5Document13 pagesCalamba Doctors' College: Digestion, Absorption, and Metabolism Lesson 5Melody DoriaNo ratings yet
- Q4 Week 3 Human Digestive System - PPTX 4thDocument47 pagesQ4 Week 3 Human Digestive System - PPTX 4thdenveralbarico143No ratings yet
- Digestive System HandoutDocument7 pagesDigestive System Handouttheodore_estradaNo ratings yet
- BiologyDocument12 pagesBiologyvidottma kolheNo ratings yet
- Digestive System 2022-23Document28 pagesDigestive System 2022-23Fady Fady100% (1)
- Lesson 4: Food and DigestionDocument8 pagesLesson 4: Food and DigestionChenul WickramasingheNo ratings yet
- Digestive GlandDocument22 pagesDigestive GlandMalinus TorquatusNo ratings yet
- Unit-V: Animal Physiology. 2Document5 pagesUnit-V: Animal Physiology. 2Sachikanta SharmaNo ratings yet
- Human & Social Biology 4th FormDocument23 pagesHuman & Social Biology 4th Formhedasdudh100% (1)
- Essay # 4. DigeDocument9 pagesEssay # 4. DigeAbdul Karim BurkiNo ratings yet
- Updated Presentation Note - 082207Document3 pagesUpdated Presentation Note - 082207Joshua AladenikaNo ratings yet
- Gastro Section CDocument11 pagesGastro Section CNadja JamilahNo ratings yet
- BiochemDocument10 pagesBiochemWaseem Abbas ShaikhNo ratings yet
- Uman Digestive System Structure and Function: DR Neelam SharmaDocument37 pagesUman Digestive System Structure and Function: DR Neelam SharmaBishal Khaidem0% (1)
- Digestion and AbsorptionDocument5 pagesDigestion and AbsorptionSheehan MathurNo ratings yet
- The Biochemistry of Digestion, Absorption and Detoxification by Prof. Dr. Hedef D. El-YassinDocument64 pagesThe Biochemistry of Digestion, Absorption and Detoxification by Prof. Dr. Hedef D. El-YassinSayhidoen CepexNo ratings yet
- Anatomy and Physiology DigestiveDocument7 pagesAnatomy and Physiology DigestiveFretzgine Lou ManuelNo ratings yet
- Biochemistry of Liver BCM 412Document20 pagesBiochemistry of Liver BCM 412onuorahu94No ratings yet
- Digestive System ScriptDocument4 pagesDigestive System ScriptNEPERESM TALCEY BORCESNo ratings yet
- Digestive System Finalppt LectureDocument30 pagesDigestive System Finalppt LectureFrancene YepesNo ratings yet
- Digestive System in Ruminents and RodentsDocument2 pagesDigestive System in Ruminents and RodentsNanaho0% (1)
- Digestion SlideDocument18 pagesDigestion SlideezebelluciNo ratings yet
- Digestive System of Poultry Avian PhysiologyDocument52 pagesDigestive System of Poultry Avian PhysiologygabrielwerneckNo ratings yet
- Digestion and Absorption - 101328Document10 pagesDigestion and Absorption - 101328Emmanuel IshiomaNo ratings yet
- Gastrointestinal System Ppt.-NewDocument117 pagesGastrointestinal System Ppt.-NewFatima Syed100% (3)
- Digestion: The Digestive SystemDocument66 pagesDigestion: The Digestive SystemTonique Gordon100% (2)
- LN Digestive SystemDocument6 pagesLN Digestive SystemCasey Dkaye MorrisNo ratings yet
- Uman Digestive System Structure and Function: Anam FarzandDocument25 pagesUman Digestive System Structure and Function: Anam FarzandTahir AzizNo ratings yet
- Digestion and NutritionDocument53 pagesDigestion and NutritionErwin Poblete AraraoNo ratings yet
- Digestive System A&p BookletDocument11 pagesDigestive System A&p Bookletnurse1990No ratings yet
- Anatomy and Physiology of Human Digestive System: The Mouth and PharynxDocument9 pagesAnatomy and Physiology of Human Digestive System: The Mouth and Pharynxitachi278No ratings yet
- Topic 6.1 - Digestion and Absorption: Stages of Processing FoodDocument5 pagesTopic 6.1 - Digestion and Absorption: Stages of Processing FoodRocio Guadalupe Lopez BlandonNo ratings yet
- Small Intestine and PancreasDocument52 pagesSmall Intestine and PancreasLutfi AuliaNo ratings yet
- The Digestion ProcessDocument7 pagesThe Digestion ProcessKim Szydney RachoNo ratings yet
- Bio C9 NotesDocument15 pagesBio C9 NotesTHEN YUE PINGNo ratings yet
- Gross Anatomy: RugaeDocument5 pagesGross Anatomy: RugaeKryza Dale Bunado BaticanNo ratings yet
- Nutrition in Humans: By: Khizar Hayat & Ade Bhai 10-ADocument40 pagesNutrition in Humans: By: Khizar Hayat & Ade Bhai 10-AWaqas TahirNo ratings yet
- 35.1 - The Digestive SystemDocument16 pages35.1 - The Digestive SystemMostafa Galal El Din100% (1)
- Git Lec 3,4Document14 pagesGit Lec 3,4Kim Sa-buNo ratings yet
- The Digestive System MODULEDocument3 pagesThe Digestive System MODULEBen Moldenhaur100% (2)
- Digestion: By: Roselita O. Natividad and Teresita G Montaño, PHD Nat. Science - Ateneo de Zamboanga UniversityDocument5 pagesDigestion: By: Roselita O. Natividad and Teresita G Montaño, PHD Nat. Science - Ateneo de Zamboanga UniversityMini BossNo ratings yet
- Human Digestion PresentationDocument35 pagesHuman Digestion PresentationYamyang Galay-BañoNo ratings yet
- Digestive SystemDocument12 pagesDigestive Systemapi-269652707No ratings yet
- Nutrition and Bioenergetics: Nature Provides A Free Lunch, But Only If We Control Our Appetite. - William RuckelshausDocument73 pagesNutrition and Bioenergetics: Nature Provides A Free Lunch, But Only If We Control Our Appetite. - William RuckelshausAndrei Allen GonzalesNo ratings yet
- Leaky Gut Syndrome Stop! - A Complete Guide To Leaky Gut Syndrome Causes, Symptoms, Treatments & A Holistic System To Eliminate LGS Naturally & PermanentlyFrom EverandLeaky Gut Syndrome Stop! - A Complete Guide To Leaky Gut Syndrome Causes, Symptoms, Treatments & A Holistic System To Eliminate LGS Naturally & PermanentlyNo ratings yet
- Movement in and Out of CellDocument19 pagesMovement in and Out of CellRand HusseinNo ratings yet
- (Spmsoalan) Soalan Biology K1 Johor 2014Document32 pages(Spmsoalan) Soalan Biology K1 Johor 2014Saidatul Atyah Mohd ApendaiNo ratings yet
- Oral Rehydration TherapyDocument7 pagesOral Rehydration TherapyBala MuruganNo ratings yet
- Fish PhysiologyDocument481 pagesFish PhysiologyfikakhanNo ratings yet
- A Level Biology NotesDocument6 pagesA Level Biology NotesMira AluedeNo ratings yet
- A Century of Exercise Physiology Key Concepts in Muscle Cell Volume RegulationDocument19 pagesA Century of Exercise Physiology Key Concepts in Muscle Cell Volume RegulationJesusNavarrete97No ratings yet
- Plant CellDocument112 pagesPlant CellAlfonso PlantillaNo ratings yet
- The Human Body: An Orientation Anatomy: Levels of Structural OrganizationDocument34 pagesThe Human Body: An Orientation Anatomy: Levels of Structural OrganizationChristian Clyde Noel JakosalemNo ratings yet
- Transport Processes and Carrier DesignDocument24 pagesTransport Processes and Carrier Designcgckamilshaikh50% (2)
- Biological Molecules/Biomolecules: O CarbohydrateDocument6 pagesBiological Molecules/Biomolecules: O CarbohydrateRamona BaculinaoNo ratings yet
- B1 RevisionDocument17 pagesB1 RevisionPierre BrocasNo ratings yet
- UntitledDocument95 pagesUntitledMohan KumarNo ratings yet
- 7 - Active-Bulk TransportDocument21 pages7 - Active-Bulk TransportCristel Jhon Ryan TagodNo ratings yet
- Cells - 4 V2 (Membranes & Transport)Document34 pagesCells - 4 V2 (Membranes & Transport)mormerodNo ratings yet
- Full Download PDF of Test Bank For Understanding Pathophysiology 6th Edition by Huether All ChapterDocument30 pagesFull Download PDF of Test Bank For Understanding Pathophysiology 6th Edition by Huether All Chapterusulliarnoob100% (4)
- Cells and Transport Study Guide MyDocument12 pagesCells and Transport Study Guide Myapi-521781723No ratings yet
- Biological Uptake Worksheet KeyDocument4 pagesBiological Uptake Worksheet KeyHanna MalinaoNo ratings yet
- Kognity Notes Bio B2Document20 pagesKognity Notes Bio B2eybasco.2001No ratings yet
- Bio Revision QuestionsDocument11 pagesBio Revision Questionsadbxjs100% (1)
- Active Passive Transport Lesson PlanDocument2 pagesActive Passive Transport Lesson Planapi-238148465100% (1)
- Mark Scheme (Results) January 2013Document9 pagesMark Scheme (Results) January 2013M7MD AshrafNo ratings yet
- O LEVEL BIOLOGY O/N 2020 PAPER 11 (5090 - w20 - QP - 11)Document20 pagesO LEVEL BIOLOGY O/N 2020 PAPER 11 (5090 - w20 - QP - 11)ImanNo ratings yet
- Answer Scheme Bio Paper 2 PPT f4 2017Document7 pagesAnswer Scheme Bio Paper 2 PPT f4 2017Norfaezah RomliNo ratings yet
- Chapter 3 Movement of Substances Across The Plasma MembraneDocument24 pagesChapter 3 Movement of Substances Across The Plasma MembraneKeith ConneryNo ratings yet
- HKALE Cytology (Cell Structure and Function) Question Paper and MSDocument18 pagesHKALE Cytology (Cell Structure and Function) Question Paper and MSNeetenin NeeteninNo ratings yet
- Bio 11 - Handout For Lectures 1-6 (Summary)Document1 pageBio 11 - Handout For Lectures 1-6 (Summary)Steffi GatdulaNo ratings yet
- Understanding Pathophysiology - ANZ Adaptation 2nd Edition by Craft - Test BankDocument37 pagesUnderstanding Pathophysiology - ANZ Adaptation 2nd Edition by Craft - Test BankalishcathrinNo ratings yet
- Biology All Master NotesDocument105 pagesBiology All Master Notesapi-260042275No ratings yet