Download as pdf or txt
You might also like
- Saunders Equine Formulary - 2nd EditionDocument511 pagesSaunders Equine Formulary - 2nd EditionSz Kinga100% (3)
- A Review of PH and OsmolarityDocument5 pagesA Review of PH and OsmolarityMuhammad Rizki SaidNo ratings yet
- 8.7 Case StudyDocument2 pages8.7 Case StudymaddisonNo ratings yet
- Efeitos Da Albumina Sobre Os EsclerosantesDocument8 pagesEfeitos Da Albumina Sobre Os EsclerosantesFelipe Ziccardi RabeloNo ratings yet
- 0049 3848 (80) 90005 5Document9 pages0049 3848 (80) 90005 5Dian NovitasariNo ratings yet
- A Novel in Vitro Assay For Anti-Inflammatory Agents Based On Stabilization of ErythrocytesDocument7 pagesA Novel in Vitro Assay For Anti-Inflammatory Agents Based On Stabilization of ErythrocytesElisabeta SavaNo ratings yet
- Man Biochemical and Pharmacological Studies With Asparaginase inDocument10 pagesMan Biochemical and Pharmacological Studies With Asparaginase inIgnacio EtchechouryNo ratings yet
- Thin-Layer Chromatography of Anticonvulsant Drugs: Clinical Chemistry April 1969Document7 pagesThin-Layer Chromatography of Anticonvulsant Drugs: Clinical Chemistry April 1969Monika NagareNo ratings yet
- Laboratory Evaluation FibrinolysisDocument4 pagesLaboratory Evaluation FibrinolysisMaritoni ReyesNo ratings yet
- 1 FullDocument9 pages1 FullFauziana NurhanisahNo ratings yet
- Role of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsDocument7 pagesRole of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsPandji 'Utuh'No ratings yet
- From The Department of Medicine, Western Reserve University, ClevelandDocument31 pagesFrom The Department of Medicine, Western Reserve University, Clevelandclarkkent2No ratings yet
- Chloremphenicol Leukemia LinkedDocument17 pagesChloremphenicol Leukemia LinkedpadbidriNo ratings yet
- Androgenic Status of Lepromatous Leprosy Patients With GynecomastiaDocument6 pagesAndrogenic Status of Lepromatous Leprosy Patients With GynecomastianadyaNo ratings yet
- A Novel, Highly Sensitive and Specific Biomarker For Niemann-Pick Type C1 DiseaseDocument8 pagesA Novel, Highly Sensitive and Specific Biomarker For Niemann-Pick Type C1 DiseaseTheodora TeddyNo ratings yet
- Fanti Et Al., 1980 (Mab To PG)Document3 pagesFanti Et Al., 1980 (Mab To PG)Farkhanda SadafNo ratings yet
- Recent Advances in Nutritional SciencesDocument4 pagesRecent Advances in Nutritional SciencesSava1988No ratings yet
- Journal Medical Research.: (New Xxviii.) Vol. XXXIII., No. January, I9I6. NoDocument14 pagesJournal Medical Research.: (New Xxviii.) Vol. XXXIII., No. January, I9I6. NoEdù Ccarhuani QuispeNo ratings yet
- Neutrophil Activation by Heme: Implications For Inflammatory ProcessesDocument7 pagesNeutrophil Activation by Heme: Implications For Inflammatory ProcessesraysellaNo ratings yet
- Petroianu 2005Document5 pagesPetroianu 2005Bilel ChefiratNo ratings yet
- 1.4 DEXTRAN-A Valuable Plasma Volume Expander. Dr.a.J.leonsins.Document6 pages1.4 DEXTRAN-A Valuable Plasma Volume Expander. Dr.a.J.leonsins.rajeevtyagi41No ratings yet
- Mechanism of Hydrogen Gas Promoted Apoptosis of Lung Adenocarcinoma A549 Cells Through XIAP and BIRC3Document12 pagesMechanism of Hydrogen Gas Promoted Apoptosis of Lung Adenocarcinoma A549 Cells Through XIAP and BIRC3FananiNo ratings yet
- Hematology 1 Lab - The Reticulocyte CountDocument17 pagesHematology 1 Lab - The Reticulocyte CountCIRILO MABBORANGNo ratings yet
- Development of A Simple Permeability Assay Method For Snake Venom Induced Vascular DamageDocument5 pagesDevelopment of A Simple Permeability Assay Method For Snake Venom Induced Vascular DamageIkhtiarNo ratings yet
- Lymphoid Crisis Myeloid Leukaemia: Blast in ChronicDocument2 pagesLymphoid Crisis Myeloid Leukaemia: Blast in ChronicThistell ThistleNo ratings yet
- 623 FullDocument1 page623 FullGabriel GlotzNo ratings yet
- Hemostasis 1Document34 pagesHemostasis 1diyoba100% (1)
- Toxins: Fibrolase: Trials and TribulationsDocument16 pagesToxins: Fibrolase: Trials and TribulationsEni PurwaeniNo ratings yet
- Cloud-Point Extraction For The Determination of The Free Fraction of Antiepileptic Drugs in Blood Plasma and SalivaDocument5 pagesCloud-Point Extraction For The Determination of The Free Fraction of Antiepileptic Drugs in Blood Plasma and SalivaDan AndonieNo ratings yet
- Chen 2009Document6 pagesChen 2009Татьяна СаенкоNo ratings yet
- APIXABANDocument14 pagesAPIXABANlorena salazarNo ratings yet
- Peripheral Blood Mononuclear Cells: Isolation, Freezing, Thawing, and CultureDocument9 pagesPeripheral Blood Mononuclear Cells: Isolation, Freezing, Thawing, and CultureGheorghe-Emilian OlteanuNo ratings yet
- Cell Free SystemDocument8 pagesCell Free SystemDuaa ShamiehNo ratings yet
- LMX 050Document19 pagesLMX 050Carolina RobinetNo ratings yet
- Plasma Homocysteine and Total Thiol Content in Patients With Exudative Age-Related Macular DegenerationDocument5 pagesPlasma Homocysteine and Total Thiol Content in Patients With Exudative Age-Related Macular DegenerationAnonymous 1jCVqQuNo ratings yet
- Non-Addictive Opium Alkaloids Selectively Induce Apoptosis in Cancer Cells Compared To Normal Cells PDFDocument8 pagesNon-Addictive Opium Alkaloids Selectively Induce Apoptosis in Cancer Cells Compared To Normal Cells PDFSantosh Kumar AgastiNo ratings yet
- 2017 BRDocument11 pages2017 BRCony GSNo ratings yet
- 2015 Article 485Document4 pages2015 Article 485davim_87No ratings yet
- Von Willebrand Factor-Cleaving Protease in Thrombotic Thrombocytopenic Purpura and The Hemolytic-Uremic SyndromeDocument7 pagesVon Willebrand Factor-Cleaving Protease in Thrombotic Thrombocytopenic Purpura and The Hemolytic-Uremic SyndromePritha BhuwapaksophonNo ratings yet
- Jurnal LisinoprilDocument7 pagesJurnal LisinoprildidiisafitriNo ratings yet
- Transfusion: Use A Low-Ionic-Strength Medium in Manual Tests For Antibody DetectionDocument6 pagesTransfusion: Use A Low-Ionic-Strength Medium in Manual Tests For Antibody DetectionRilind MuratiNo ratings yet
- Use of Peripheral Blood Leukocytes in The Study of Cell-Mediated Immunity in Bovine Anaplasmosis - A ReviewDocument5 pagesUse of Peripheral Blood Leukocytes in The Study of Cell-Mediated Immunity in Bovine Anaplasmosis - A Reviewyudhi arjentiniaNo ratings yet
- Erythrocytes As Novel Drug Delivery SystemDocument18 pagesErythrocytes As Novel Drug Delivery SystemMayson BaliNo ratings yet
- Ijcem0008 10558Document10 pagesIjcem0008 10558Alber AvendañoNo ratings yet
- Artificial Blood and Blood SubstitiuteDocument34 pagesArtificial Blood and Blood SubstitiuteASHISH RANJANNo ratings yet
- Self-Powered Sensor For Naked-Eye Detection of Serum TrypsinDocument4 pagesSelf-Powered Sensor For Naked-Eye Detection of Serum TrypsinCons MirandaNo ratings yet
- Wang 2015Document14 pagesWang 2015Manuel Esquivel OrtegaNo ratings yet
- Pharmacokinetics and Efficacy of Chlorpheniramine in ChildrenDocument6 pagesPharmacokinetics and Efficacy of Chlorpheniramine in ChildrenlilingNo ratings yet
- Aac00051 0112Document9 pagesAac00051 0112Rocio AlejandraNo ratings yet
- Hematin InhibitorDocument5 pagesHematin InhibitorApriliana Wulandari SyafitriNo ratings yet
- Acute B-Lymphoid Leukemia in A Mare: A Case Report: B. Bezdekova, M. Faldyna, O. Zapletal, P. JahnDocument7 pagesAcute B-Lymphoid Leukemia in A Mare: A Case Report: B. Bezdekova, M. Faldyna, O. Zapletal, P. JahnRaisis Farah DzakiyyahNo ratings yet
- Bencimidazoles Anti CáncerDocument10 pagesBencimidazoles Anti CáncerEylan Goitia De GonzálezNo ratings yet
- Acetaminophen Increases Aldosterone Secretion While SuppressingDocument7 pagesAcetaminophen Increases Aldosterone Secretion While SuppressingKhánh Nguyễn NgọcNo ratings yet
- Hairy Cell Leukemia: Functional, Immunologic, Kinetic, and Ultrastructural CharacterizationDocument13 pagesHairy Cell Leukemia: Functional, Immunologic, Kinetic, and Ultrastructural CharacterizationAna Rubí LópezNo ratings yet
- Western Blot AnalysisDocument5 pagesWestern Blot AnalysiskapilphysioNo ratings yet
- Observations Hemoglobin: On The Minor Basic Component IN Blood of Normal Individuals and Patients With ThalassemiaDocument11 pagesObservations Hemoglobin: On The Minor Basic Component IN Blood of Normal Individuals and Patients With ThalassemiaSudhit SethiNo ratings yet
- 375 Full PDFDocument5 pages375 Full PDFDiky MudhakirNo ratings yet
- PselectinDocument7 pagesPselectinAstarag DasNo ratings yet
- The Spectrum of Amniotic Fluid Embolism: Is Intralipid the solution ?From EverandThe Spectrum of Amniotic Fluid Embolism: Is Intralipid the solution ?No ratings yet
- Pheochromocytomas, Paragangliomas and Disorders of the Sympathoadrenal System: Clinical Features, Diagnosis and ManagementFrom EverandPheochromocytomas, Paragangliomas and Disorders of the Sympathoadrenal System: Clinical Features, Diagnosis and ManagementLewis LandsbergNo ratings yet
- Further Studies On The Erythrocytes Anti Inflammatory AssayDocument6 pagesFurther Studies On The Erythrocytes Anti Inflammatory AssayElisabeta SavaNo ratings yet
- In Vitro Effects of Non-Steroidal Anti-Inflammatory Drugs (NAIFD)Document6 pagesIn Vitro Effects of Non-Steroidal Anti-Inflammatory Drugs (NAIFD)Elisabeta SavaNo ratings yet
- Handbook of Experimental Pharmacology Anti-Inflammatory-DrugsDocument981 pagesHandbook of Experimental Pharmacology Anti-Inflammatory-DrugsElisabeta SavaNo ratings yet
- A Novel in Vitro Assay For Anti-Inflammatory Agents Based On Stabilization of ErythrocytesDocument7 pagesA Novel in Vitro Assay For Anti-Inflammatory Agents Based On Stabilization of ErythrocytesElisabeta SavaNo ratings yet
- NS 1 Quiz 7 Sas 19 23Document15 pagesNS 1 Quiz 7 Sas 19 23it's me ANGELANo ratings yet
- PMLS1 Lesson 3Document8 pagesPMLS1 Lesson 3John Daniel AriasNo ratings yet
- SAGM Blood For Neonatal Large Volume TransfusionDocument4 pagesSAGM Blood For Neonatal Large Volume TransfusionMerhan FoudaNo ratings yet
- Module 2 PPT NOTES HematopoiesisDocument8 pagesModule 2 PPT NOTES HematopoiesisMohammad MasacalNo ratings yet
- Hematological Anatomy, Physiology and Assessment: This Course Has Been Awarded One (1.0) Contact Hour. July 30, 2019Document19 pagesHematological Anatomy, Physiology and Assessment: This Course Has Been Awarded One (1.0) Contact Hour. July 30, 2019Mill Jan CruzNo ratings yet
- Equipment Specification SL-1Document21 pagesEquipment Specification SL-1Ibrahim KamaraNo ratings yet
- Blood 2Document13 pagesBlood 2abbaslafeNo ratings yet
- Blood Homework C COMPLETEDocument20 pagesBlood Homework C COMPLETEbvalery97No ratings yet
- The Toxicity Effect of Monocrotophos 36% E. C On The Haematology, LabeoDocument4 pagesThe Toxicity Effect of Monocrotophos 36% E. C On The Haematology, LabeoTamizhazhaganNo ratings yet
- Jurnal - Komponen DarahDocument6 pagesJurnal - Komponen Darahvivi maykasariNo ratings yet
- Circulatory SystemDocument7 pagesCirculatory SystemWisdom DzombeNo ratings yet
- Common Medical AbbreviationsDocument4 pagesCommon Medical AbbreviationsApril Joy de LimaNo ratings yet
- Forensic Medicine: Forensic Medicine - The Science That Deals With The Application ofDocument16 pagesForensic Medicine: Forensic Medicine - The Science That Deals With The Application ofFull GamingClipsNo ratings yet
- Diagnostic Approach To Anemia in Adults - UpToDateDocument76 pagesDiagnostic Approach To Anemia in Adults - UpToDateanweridrisejazNo ratings yet
- Protection of Oxidative Damage by Aqueous Extract.Document14 pagesProtection of Oxidative Damage by Aqueous Extract.Phan Đình HuyNo ratings yet
- Chapter 11Document11 pagesChapter 11christian anchetaNo ratings yet
- Fchem 10 905256Document16 pagesFchem 10 905256os krishnaNo ratings yet
- Laporan Sel Darah Merah (Eritrosit)Document8 pagesLaporan Sel Darah Merah (Eritrosit)RindiWirantikaNo ratings yet
- Definitions: CLS 422 Clinical Immunohematology I Absorption and ElutionDocument6 pagesDefinitions: CLS 422 Clinical Immunohematology I Absorption and ElutionTrang HuynhNo ratings yet
- JAYPEE Handbook of Blood Banking - Transfusion Medicine, 2006Document366 pagesJAYPEE Handbook of Blood Banking - Transfusion Medicine, 2006Dwi della yaumil qitmiaNo ratings yet
- Biology - Human Transport NotesDocument4 pagesBiology - Human Transport Notesacsbr4science133% (3)
- Assessment Nursing Diagnosis Background Knowledge Planning Intervention Rationale EvaluationDocument5 pagesAssessment Nursing Diagnosis Background Knowledge Planning Intervention Rationale EvaluationJackRolunaBernalesNo ratings yet
- Topics of This Lecture: RBC: - Structural Characteristics - Hemoglobin - Erythropoiesis - Erythrocytes DestructionDocument23 pagesTopics of This Lecture: RBC: - Structural Characteristics - Hemoglobin - Erythropoiesis - Erythrocytes DestructionHashim GhazoNo ratings yet
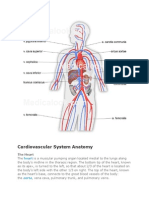
- Cardiovascular System AnatomyDocument16 pagesCardiovascular System AnatomyJona Addatu0% (1)
- Hematology: Michele L. Miller, RN, MSN-NEDocument33 pagesHematology: Michele L. Miller, RN, MSN-NEniekro35100% (1)
- 82-P01.91.300027-04 KT-6400英文说明书Document132 pages82-P01.91.300027-04 KT-6400英文说明书郑伟健No ratings yet
- DNeasy Blood & Tissue HandbookDocument62 pagesDNeasy Blood & Tissue HandbookPeter Hong Leong CheahNo ratings yet
- 3600+ Review Questions For Anatomy & Physiology Volume 2 - (4th Edition) PDFDocument151 pages3600+ Review Questions For Anatomy & Physiology Volume 2 - (4th Edition) PDFMauzoom Ali100% (2)