
Descarboxilación Del Piruvato
Descarboxilación Del Piruvato
You might also like
- Inorganic Chemistry 5Th Edition Miessler Solutions Manual Full Chapter PDFDocument37 pagesInorganic Chemistry 5Th Edition Miessler Solutions Manual Full Chapter PDFgregorythurv0e81% (16)
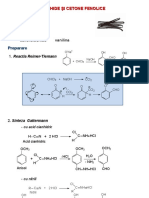
- Aldehide Şi Cetone FenoliceDocument9 pagesAldehide Şi Cetone FenoliceMarinelaNo ratings yet
- Central Dogma WebquestDocument4 pagesCentral Dogma Webquestapi-446838554No ratings yet
- Pyruvate Dehydrogenase Complex: BiochemistryDocument24 pagesPyruvate Dehydrogenase Complex: BiochemistrySANDIP PATILNo ratings yet
- NEPHAR 305 Metabolism - 12Document61 pagesNEPHAR 305 Metabolism - 12Ra'fat RaheemNo ratings yet
- Linked ListDocument3 pagesLinked ListpragatiraichanNo ratings yet
- Amine Unit Materials and Corrosion (Updated)Document36 pagesAmine Unit Materials and Corrosion (Updated)harrinsonfNo ratings yet
- Matriculation Chemistry (Amino Acids) Part 2Document10 pagesMatriculation Chemistry (Amino Acids) Part 2ridwanNo ratings yet
- CH12 Unit04 AOSRA02Document7 pagesCH12 Unit04 AOSRA02PyNo ratings yet
- Enzyme&RegulationDocument51 pagesEnzyme&Regulationfaisal wisnuajiNo ratings yet
- 1 An Observation - The Krebs CycleDocument33 pages1 An Observation - The Krebs CycleOmbrog JustinNo ratings yet
- MANASVIIDocument20 pagesMANASVIICyber MagicNo ratings yet
- Chela Tesch Elating Agents 2017Document3 pagesChela Tesch Elating Agents 2017Sharin Bin Ab GhaniNo ratings yet
- Pyrimidine de Novo Synthesis RecapDocument3 pagesPyrimidine de Novo Synthesis RecapMacarie CosminaNo ratings yet
- Nucleophilic SubstitutionDocument22 pagesNucleophilic Substitutionnishitagupta790No ratings yet
- Lecture 15a-CatalysisDocument52 pagesLecture 15a-CatalysisNagarjuna VuchuruNo ratings yet
- Aldehydes and KetonesDocument41 pagesAldehydes and KetonesJerome DimaanoNo ratings yet
- 5 Aldehydes and Ketones-Structure and PreparationDocument41 pages5 Aldehydes and Ketones-Structure and PreparationKeshav JoshiNo ratings yet
- Aldehyde and KetonesDocument41 pagesAldehyde and KetonesJerome DimaanoNo ratings yet
- 100S120 CS19L01Document38 pages100S120 CS19L01b101112154No ratings yet
- TCA CycleDocument35 pagesTCA CycleSantino MajokNo ratings yet
- Ald&Ketone IDocument41 pagesAld&Ketone IHarsha Y MNo ratings yet
- Biosíntesis de Purinas y PirimidinasDocument18 pagesBiosíntesis de Purinas y PirimidinasXimenaNo ratings yet
- Fatty Acid Oxidation: Molecular Biochemistry IIDocument43 pagesFatty Acid Oxidation: Molecular Biochemistry IIMurtaza Hajoori100% (1)
- Reductions PPT 29-08-2020Document12 pagesReductions PPT 29-08-2020jkc collegeNo ratings yet
- Solution of Chemistry HSSC-II (3rd Set)Document11 pagesSolution of Chemistry HSSC-II (3rd Set)Ujala ShahidNo ratings yet
- Karbohidrat KuliahDocument38 pagesKarbohidrat KuliahMOCHILNo ratings yet
- Metabolisme ProteinDocument28 pagesMetabolisme ProteinFerlisan TabanciNo ratings yet
- OXIDATIONS FinalDocument9 pagesOXIDATIONS Finalgamer boomerNo ratings yet
- AGE and KidneyDocument14 pagesAGE and KidneyKarina ThiemeNo ratings yet
- Fatty Acid Oxidation: Molecular Biochemistry IIDocument39 pagesFatty Acid Oxidation: Molecular Biochemistry IIDozdi100% (1)
- TutorialDocument27 pagesTutorialSiti NuraqidahNo ratings yet
- Aldehydes, Ketones and Carboxylic Acids _ Short NotesDocument2 pagesAldehydes, Ketones and Carboxylic Acids _ Short Notesrajvirg100No ratings yet
- Aldehydes, Ketones and Carboxylic Acids - Short NotesDocument2 pagesAldehydes, Ketones and Carboxylic Acids - Short NotesAyush PalNo ratings yet
- Preparation of MaleimideDocument11 pagesPreparation of MaleimideDotsha Raheem100% (1)
- Comp Carbon Ilic IDocument22 pagesComp Carbon Ilic IAndreea Andreea S100% (1)
- Carbohydrates - Copy New SimplifiedDocument74 pagesCarbohydrates - Copy New SimplifiedWnwixiejwis Usisjdx8xjwiNo ratings yet
- Heter 0Document22 pagesHeter 0Lot AdewumilotNo ratings yet
- Carbohydrates PresentationDocument60 pagesCarbohydrates PresentationRaagas, ChristianNo ratings yet
- Requirement: Unsaturated Monomer: Addition PolymerizationDocument24 pagesRequirement: Unsaturated Monomer: Addition PolymerizationLester John VeraNo ratings yet
- GLUCIDE-lipide-structuri-1-2015 - ENGDocument53 pagesGLUCIDE-lipide-structuri-1-2015 - ENGCodrin FodorNo ratings yet
- Chapter FiveDocument24 pagesChapter FiveAbhilasha VashisthaNo ratings yet
- C - Sol - Ch-26 - Aldehydes Ketones and Carboxylic AcidsDocument19 pagesC - Sol - Ch-26 - Aldehydes Ketones and Carboxylic AcidsHimanshi ChahalNo ratings yet
- N, N-Dimethylaniline + Nano: L D D D D D D DDocument10 pagesN, N-Dimethylaniline + Nano: L D D D D D D DNene TanNo ratings yet
- NH Chemistry All 2019-DQP-SpellcheckonDocument65 pagesNH Chemistry All 2019-DQP-SpellcheckonJohn TechNo ratings yet
- Synthesis of Drug 22Document44 pagesSynthesis of Drug 22sdfsy sdvdsvNo ratings yet
- Lec 4 Kerbs - CycleDocument41 pagesLec 4 Kerbs - Cyclemicklemagdy50No ratings yet
- Sintesis PeptidaDocument19 pagesSintesis PeptidaHasna NoerNo ratings yet
- KarbohidratDocument62 pagesKarbohidratfachira auliaNo ratings yet
- ReductionDocument36 pagesReductionSayed Newaj ChowdhuryNo ratings yet
- Biomolecules: Section (A) : CarbohydratesDocument29 pagesBiomolecules: Section (A) : CarbohydratesUma KasyapNo ratings yet
- Ultima Named ReactionsDocument88 pagesUltima Named ReactionsKrish RawatNo ratings yet
- 4L OxidationsDocument16 pages4L OxidationsCarlos Javier Orellana OrtizNo ratings yet
- 11 - Miscellaneous ReactionDocument4 pages11 - Miscellaneous ReactionMaitreya DasNo ratings yet
- Chapter 11: Nucleophilic Substitution and Elimination: Ho O PCL Oh Ho ODocument19 pagesChapter 11: Nucleophilic Substitution and Elimination: Ho O PCL Oh Ho OVENUGOPALARAONo ratings yet
- Chemical RXN - Aldehydes, KetoneDocument11 pagesChemical RXN - Aldehydes, Ketoneudhayadeepak60No ratings yet
- Chapter 5 HydrocarbonDocument25 pagesChapter 5 Hydrocarbonmeshal retteryNo ratings yet
- Lipida: Indah Saraswati, M. SCDocument55 pagesLipida: Indah Saraswati, M. SCPutriNo ratings yet
- ლექცია 8. პოლიმერებისათვის საჭირო საწყისი მონომერების მიღების და შუალედური პროდუქტების მიღებაDocument30 pagesლექცია 8. პოლიმერებისათვის საჭირო საწყისი მონომერების მიღების და შუალედური პროდუქტების მიღებაtoduamaiko8No ratings yet
- Advanced Biochemistry: The Krebs CycleDocument11 pagesAdvanced Biochemistry: The Krebs CycleMaritsa PerHerNo ratings yet
- BLOSUM MatricesDocument18 pagesBLOSUM Matricesshushantobanik77No ratings yet
- Molecular Cell Biology Lodish 6th Edition Test BankDocument6 pagesMolecular Cell Biology Lodish 6th Edition Test Bankgenevievedoilf2opNo ratings yet
- Jasdeep Singh, PHD, Nasreen Z. Ehtesham, Syed Asad Rahman, PHD, Seyed E. Hasnain, PHDDocument8 pagesJasdeep Singh, PHD, Nasreen Z. Ehtesham, Syed Asad Rahman, PHD, Seyed E. Hasnain, PHDMuhammad Riza DarmawanNo ratings yet
- Genetic EngineeringDocument35 pagesGenetic Engineeringjosh321No ratings yet
- 8 BiodiversityDocument10 pages8 BiodiversityAra Villamor YusopNo ratings yet
- Progress Report of MR V.K.singh Information OfficerDocument2 pagesProgress Report of MR V.K.singh Information OfficerVinay SinghNo ratings yet
- Bioinformatics NoteDocument7 pagesBioinformatics NoteNITI SHAHNo ratings yet
- MetabolomicsDocument5 pagesMetabolomicsFitri Nur AiniNo ratings yet
- Session 1: The Role of Plant Breeding in Meeting The Multiple Challenges of A Fast-Changing WorldDocument72 pagesSession 1: The Role of Plant Breeding in Meeting The Multiple Challenges of A Fast-Changing WorldMourad Taibi100% (2)
- Assignment of Pathology 401: Submitted byDocument5 pagesAssignment of Pathology 401: Submitted byaymen gulzarNo ratings yet
- Feschotte 2007Document38 pagesFeschotte 2007Axel GómezNo ratings yet
- KB FileDocument18 pagesKB FileKaushik BanikNo ratings yet
- Transgenic AnimaliaDocument4 pagesTransgenic AnimaliaGalino Julia Cristine A.No ratings yet
- Zoology - 04-Biomolecule 2023 NotesDocument18 pagesZoology - 04-Biomolecule 2023 NotesSquad 4 GamingNo ratings yet
- Quatum DotsDocument9 pagesQuatum DotssherlyNo ratings yet
- Microbiology Lecture Exam Unit #2 Study Guide Essential Biology and Survey of The Microbial WorldDocument6 pagesMicrobiology Lecture Exam Unit #2 Study Guide Essential Biology and Survey of The Microbial WorldTatum MastermanNo ratings yet
- Patofisiologi Stroke Iskemik IbnDocument14 pagesPatofisiologi Stroke Iskemik Ibnashwin kumarNo ratings yet
- Biology Module 1 Lesson 1 Genetic EngineeringDocument11 pagesBiology Module 1 Lesson 1 Genetic EngineeringHannah Joy D. AROSTIQUENo ratings yet
- Cross Priming AmplificationDocument7 pagesCross Priming Amplificationmateus_laguardiaNo ratings yet
- June 2015 QP - Unit 5 WJEC Biology A-LevelDocument24 pagesJune 2015 QP - Unit 5 WJEC Biology A-LevelNonuNo ratings yet
- Lab 9, RFLPDocument5 pagesLab 9, RFLPPatryk RogNo ratings yet
- II.D. How Organisms Obtain and Utilize EnergyDocument19 pagesII.D. How Organisms Obtain and Utilize EnergyDarwin Nool100% (2)
- Lab 9 - 022-023Document4 pagesLab 9 - 022-023Noor JanahiNo ratings yet
- Manning Valley Anglican College Assessment Notification AdviceDocument5 pagesManning Valley Anglican College Assessment Notification AdviceADNo ratings yet
- Traditonal Vs Modern Biotechnology PDFDocument4 pagesTraditonal Vs Modern Biotechnology PDFKc Lagran50% (2)
- Small Animal Clinical NutritionDocument6 pagesSmall Animal Clinical NutritionJairo Pereira100% (1)
- Exercise 12Document7 pagesExercise 12Lennon DavalosNo ratings yet
- Polymerase Chain Reac6on PCR: What To Do Today?Document7 pagesPolymerase Chain Reac6on PCR: What To Do Today?Juliana MaltaNo ratings yet
- Biochemistry Revalida 2021Document30 pagesBiochemistry Revalida 2021RM DulawanNo ratings yet

























































































Download as pdf or txt
You might also like
- Inorganic Chemistry 5Th Edition Miessler Solutions Manual Full Chapter PDFDocument37 pagesInorganic Chemistry 5Th Edition Miessler Solutions Manual Full Chapter PDFgregorythurv0e81% (16)
- Aldehide Şi Cetone FenoliceDocument9 pagesAldehide Şi Cetone FenoliceMarinelaNo ratings yet
- Central Dogma WebquestDocument4 pagesCentral Dogma Webquestapi-446838554No ratings yet
- Pyruvate Dehydrogenase Complex: BiochemistryDocument24 pagesPyruvate Dehydrogenase Complex: BiochemistrySANDIP PATILNo ratings yet
- NEPHAR 305 Metabolism - 12Document61 pagesNEPHAR 305 Metabolism - 12Ra'fat RaheemNo ratings yet
- Linked ListDocument3 pagesLinked ListpragatiraichanNo ratings yet
- Amine Unit Materials and Corrosion (Updated)Document36 pagesAmine Unit Materials and Corrosion (Updated)harrinsonfNo ratings yet
- Matriculation Chemistry (Amino Acids) Part 2Document10 pagesMatriculation Chemistry (Amino Acids) Part 2ridwanNo ratings yet
- CH12 Unit04 AOSRA02Document7 pagesCH12 Unit04 AOSRA02PyNo ratings yet
- Enzyme&RegulationDocument51 pagesEnzyme&Regulationfaisal wisnuajiNo ratings yet
- 1 An Observation - The Krebs CycleDocument33 pages1 An Observation - The Krebs CycleOmbrog JustinNo ratings yet
- MANASVIIDocument20 pagesMANASVIICyber MagicNo ratings yet
- Chela Tesch Elating Agents 2017Document3 pagesChela Tesch Elating Agents 2017Sharin Bin Ab GhaniNo ratings yet
- Pyrimidine de Novo Synthesis RecapDocument3 pagesPyrimidine de Novo Synthesis RecapMacarie CosminaNo ratings yet
- Nucleophilic SubstitutionDocument22 pagesNucleophilic Substitutionnishitagupta790No ratings yet
- Lecture 15a-CatalysisDocument52 pagesLecture 15a-CatalysisNagarjuna VuchuruNo ratings yet
- Aldehydes and KetonesDocument41 pagesAldehydes and KetonesJerome DimaanoNo ratings yet
- 5 Aldehydes and Ketones-Structure and PreparationDocument41 pages5 Aldehydes and Ketones-Structure and PreparationKeshav JoshiNo ratings yet
- Aldehyde and KetonesDocument41 pagesAldehyde and KetonesJerome DimaanoNo ratings yet
- 100S120 CS19L01Document38 pages100S120 CS19L01b101112154No ratings yet
- TCA CycleDocument35 pagesTCA CycleSantino MajokNo ratings yet
- Ald&Ketone IDocument41 pagesAld&Ketone IHarsha Y MNo ratings yet
- Biosíntesis de Purinas y PirimidinasDocument18 pagesBiosíntesis de Purinas y PirimidinasXimenaNo ratings yet
- Fatty Acid Oxidation: Molecular Biochemistry IIDocument43 pagesFatty Acid Oxidation: Molecular Biochemistry IIMurtaza Hajoori100% (1)
- Reductions PPT 29-08-2020Document12 pagesReductions PPT 29-08-2020jkc collegeNo ratings yet
- Solution of Chemistry HSSC-II (3rd Set)Document11 pagesSolution of Chemistry HSSC-II (3rd Set)Ujala ShahidNo ratings yet
- Karbohidrat KuliahDocument38 pagesKarbohidrat KuliahMOCHILNo ratings yet
- Metabolisme ProteinDocument28 pagesMetabolisme ProteinFerlisan TabanciNo ratings yet
- OXIDATIONS FinalDocument9 pagesOXIDATIONS Finalgamer boomerNo ratings yet
- AGE and KidneyDocument14 pagesAGE and KidneyKarina ThiemeNo ratings yet
- Fatty Acid Oxidation: Molecular Biochemistry IIDocument39 pagesFatty Acid Oxidation: Molecular Biochemistry IIDozdi100% (1)
- TutorialDocument27 pagesTutorialSiti NuraqidahNo ratings yet
- Aldehydes, Ketones and Carboxylic Acids _ Short NotesDocument2 pagesAldehydes, Ketones and Carboxylic Acids _ Short Notesrajvirg100No ratings yet
- Aldehydes, Ketones and Carboxylic Acids - Short NotesDocument2 pagesAldehydes, Ketones and Carboxylic Acids - Short NotesAyush PalNo ratings yet
- Preparation of MaleimideDocument11 pagesPreparation of MaleimideDotsha Raheem100% (1)
- Comp Carbon Ilic IDocument22 pagesComp Carbon Ilic IAndreea Andreea S100% (1)
- Carbohydrates - Copy New SimplifiedDocument74 pagesCarbohydrates - Copy New SimplifiedWnwixiejwis Usisjdx8xjwiNo ratings yet
- Heter 0Document22 pagesHeter 0Lot AdewumilotNo ratings yet
- Carbohydrates PresentationDocument60 pagesCarbohydrates PresentationRaagas, ChristianNo ratings yet
- Requirement: Unsaturated Monomer: Addition PolymerizationDocument24 pagesRequirement: Unsaturated Monomer: Addition PolymerizationLester John VeraNo ratings yet
- GLUCIDE-lipide-structuri-1-2015 - ENGDocument53 pagesGLUCIDE-lipide-structuri-1-2015 - ENGCodrin FodorNo ratings yet
- Chapter FiveDocument24 pagesChapter FiveAbhilasha VashisthaNo ratings yet
- C - Sol - Ch-26 - Aldehydes Ketones and Carboxylic AcidsDocument19 pagesC - Sol - Ch-26 - Aldehydes Ketones and Carboxylic AcidsHimanshi ChahalNo ratings yet
- N, N-Dimethylaniline + Nano: L D D D D D D DDocument10 pagesN, N-Dimethylaniline + Nano: L D D D D D D DNene TanNo ratings yet
- NH Chemistry All 2019-DQP-SpellcheckonDocument65 pagesNH Chemistry All 2019-DQP-SpellcheckonJohn TechNo ratings yet
- Synthesis of Drug 22Document44 pagesSynthesis of Drug 22sdfsy sdvdsvNo ratings yet
- Lec 4 Kerbs - CycleDocument41 pagesLec 4 Kerbs - Cyclemicklemagdy50No ratings yet
- Sintesis PeptidaDocument19 pagesSintesis PeptidaHasna NoerNo ratings yet
- KarbohidratDocument62 pagesKarbohidratfachira auliaNo ratings yet
- ReductionDocument36 pagesReductionSayed Newaj ChowdhuryNo ratings yet
- Biomolecules: Section (A) : CarbohydratesDocument29 pagesBiomolecules: Section (A) : CarbohydratesUma KasyapNo ratings yet
- Ultima Named ReactionsDocument88 pagesUltima Named ReactionsKrish RawatNo ratings yet
- 4L OxidationsDocument16 pages4L OxidationsCarlos Javier Orellana OrtizNo ratings yet
- 11 - Miscellaneous ReactionDocument4 pages11 - Miscellaneous ReactionMaitreya DasNo ratings yet
- Chapter 11: Nucleophilic Substitution and Elimination: Ho O PCL Oh Ho ODocument19 pagesChapter 11: Nucleophilic Substitution and Elimination: Ho O PCL Oh Ho OVENUGOPALARAONo ratings yet
- Chemical RXN - Aldehydes, KetoneDocument11 pagesChemical RXN - Aldehydes, Ketoneudhayadeepak60No ratings yet
- Chapter 5 HydrocarbonDocument25 pagesChapter 5 Hydrocarbonmeshal retteryNo ratings yet
- Lipida: Indah Saraswati, M. SCDocument55 pagesLipida: Indah Saraswati, M. SCPutriNo ratings yet
- ლექცია 8. პოლიმერებისათვის საჭირო საწყისი მონომერების მიღების და შუალედური პროდუქტების მიღებაDocument30 pagesლექცია 8. პოლიმერებისათვის საჭირო საწყისი მონომერების მიღების და შუალედური პროდუქტების მიღებაtoduamaiko8No ratings yet
- Advanced Biochemistry: The Krebs CycleDocument11 pagesAdvanced Biochemistry: The Krebs CycleMaritsa PerHerNo ratings yet
- BLOSUM MatricesDocument18 pagesBLOSUM Matricesshushantobanik77No ratings yet
- Molecular Cell Biology Lodish 6th Edition Test BankDocument6 pagesMolecular Cell Biology Lodish 6th Edition Test Bankgenevievedoilf2opNo ratings yet
- Jasdeep Singh, PHD, Nasreen Z. Ehtesham, Syed Asad Rahman, PHD, Seyed E. Hasnain, PHDDocument8 pagesJasdeep Singh, PHD, Nasreen Z. Ehtesham, Syed Asad Rahman, PHD, Seyed E. Hasnain, PHDMuhammad Riza DarmawanNo ratings yet
- Genetic EngineeringDocument35 pagesGenetic Engineeringjosh321No ratings yet
- 8 BiodiversityDocument10 pages8 BiodiversityAra Villamor YusopNo ratings yet
- Progress Report of MR V.K.singh Information OfficerDocument2 pagesProgress Report of MR V.K.singh Information OfficerVinay SinghNo ratings yet
- Bioinformatics NoteDocument7 pagesBioinformatics NoteNITI SHAHNo ratings yet
- MetabolomicsDocument5 pagesMetabolomicsFitri Nur AiniNo ratings yet
- Session 1: The Role of Plant Breeding in Meeting The Multiple Challenges of A Fast-Changing WorldDocument72 pagesSession 1: The Role of Plant Breeding in Meeting The Multiple Challenges of A Fast-Changing WorldMourad Taibi100% (2)
- Assignment of Pathology 401: Submitted byDocument5 pagesAssignment of Pathology 401: Submitted byaymen gulzarNo ratings yet
- Feschotte 2007Document38 pagesFeschotte 2007Axel GómezNo ratings yet
- KB FileDocument18 pagesKB FileKaushik BanikNo ratings yet
- Transgenic AnimaliaDocument4 pagesTransgenic AnimaliaGalino Julia Cristine A.No ratings yet
- Zoology - 04-Biomolecule 2023 NotesDocument18 pagesZoology - 04-Biomolecule 2023 NotesSquad 4 GamingNo ratings yet
- Quatum DotsDocument9 pagesQuatum DotssherlyNo ratings yet
- Microbiology Lecture Exam Unit #2 Study Guide Essential Biology and Survey of The Microbial WorldDocument6 pagesMicrobiology Lecture Exam Unit #2 Study Guide Essential Biology and Survey of The Microbial WorldTatum MastermanNo ratings yet
- Patofisiologi Stroke Iskemik IbnDocument14 pagesPatofisiologi Stroke Iskemik Ibnashwin kumarNo ratings yet
- Biology Module 1 Lesson 1 Genetic EngineeringDocument11 pagesBiology Module 1 Lesson 1 Genetic EngineeringHannah Joy D. AROSTIQUENo ratings yet
- Cross Priming AmplificationDocument7 pagesCross Priming Amplificationmateus_laguardiaNo ratings yet
- June 2015 QP - Unit 5 WJEC Biology A-LevelDocument24 pagesJune 2015 QP - Unit 5 WJEC Biology A-LevelNonuNo ratings yet
- Lab 9, RFLPDocument5 pagesLab 9, RFLPPatryk RogNo ratings yet
- II.D. How Organisms Obtain and Utilize EnergyDocument19 pagesII.D. How Organisms Obtain and Utilize EnergyDarwin Nool100% (2)
- Lab 9 - 022-023Document4 pagesLab 9 - 022-023Noor JanahiNo ratings yet
- Manning Valley Anglican College Assessment Notification AdviceDocument5 pagesManning Valley Anglican College Assessment Notification AdviceADNo ratings yet
- Traditonal Vs Modern Biotechnology PDFDocument4 pagesTraditonal Vs Modern Biotechnology PDFKc Lagran50% (2)
- Small Animal Clinical NutritionDocument6 pagesSmall Animal Clinical NutritionJairo Pereira100% (1)
- Exercise 12Document7 pagesExercise 12Lennon DavalosNo ratings yet
- Polymerase Chain Reac6on PCR: What To Do Today?Document7 pagesPolymerase Chain Reac6on PCR: What To Do Today?Juliana MaltaNo ratings yet
- Biochemistry Revalida 2021Document30 pagesBiochemistry Revalida 2021RM DulawanNo ratings yet