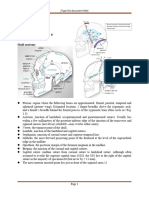
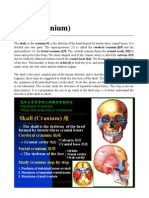
Download as pdf or txt
You might also like
- Anatomy and Physiology 6th Edition Marieb Solutions Manual Full Chapter PDFDocument41 pagesAnatomy and Physiology 6th Edition Marieb Solutions Manual Full Chapter PDFserenafinnodx100% (11)
- Laboratory Manual For Anatomy and Physiology 6th Edition Marieb Solutions ManualDocument7 pagesLaboratory Manual For Anatomy and Physiology 6th Edition Marieb Solutions Manualhanhgiaoghhr100% (37)
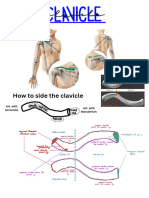
- Chapter 8 Lab Assignment - Appendicular Skeleton LabelingDocument6 pagesChapter 8 Lab Assignment - Appendicular Skeleton Labelingadriana blanco galianoNo ratings yet
- Systematic Osteology 2020Document20 pagesSystematic Osteology 2020KalangiIrushikaNo ratings yet
- LAB_Parts of the human brainDocument5 pagesLAB_Parts of the human brainNathaniel GuirhemNo ratings yet
- Chapter07 TheHead&Neck 150 165 PDFDocument16 pagesChapter07 TheHead&Neck 150 165 PDFPopa AlexandraNo ratings yet
- All Bony LandmarkDocument34 pagesAll Bony Landmarkvickytang22072002No ratings yet
- Neuro Lab (Spinal Cord+Telencephalon)2024Document19 pagesNeuro Lab (Spinal Cord+Telencephalon)2024Maneesri NakvijetreNo ratings yet
- Anatomy of The SkullDocument29 pagesAnatomy of The Skullaya abdullahNo ratings yet
- Lab 7 Pectoral Girdle 2Document4 pagesLab 7 Pectoral Girdle 2Bechris BambiNo ratings yet
- Integrative Functions of The Nervous SystemDocument18 pagesIntegrative Functions of The Nervous Systempuchio100% (1)
- Anatomy and Physiology 6th Edition Marieb Solutions ManualDocument20 pagesAnatomy and Physiology 6th Edition Marieb Solutions Manualstephenthanh1huo100% (33)
- Lec.2 H&N Anatomy, BuraqDocument55 pagesLec.2 H&N Anatomy, BuraqAkramNo ratings yet
- Anatomy Lect of Skull 2Document36 pagesAnatomy Lect of Skull 2sanullah123khan.13No ratings yet
- 1anatomy of The Nervous System Part I PDFDocument79 pages1anatomy of The Nervous System Part I PDFOmar AlemánNo ratings yet
- Virtual Laboratory Activity Worksheet On Central Nervous SystemDocument6 pagesVirtual Laboratory Activity Worksheet On Central Nervous SystemAngela ParaisoNo ratings yet
- l15 CerebrumDocument31 pagesl15 CerebrumDr. Pinki RaiNo ratings yet
- Johns 2014Document21 pagesJohns 2014SAYDE ILEANA NAJERA DOMINGUEZNo ratings yet
- NEUROANATOMYDocument34 pagesNEUROANATOMYRavanno Fanizza HarahapNo ratings yet
- Yyyyycrash Course Anatomy - Head & NeckDocument52 pagesYyyyycrash Course Anatomy - Head & Neck黃芳昌No ratings yet
- Anatomy and Physiology 6th Edition Marieb Solutions Manual instant download all chapterDocument42 pagesAnatomy and Physiology 6th Edition Marieb Solutions Manual instant download all chapterbanguepengli100% (3)
- 03 Meninges, Neuron, NMJ, CSF (Students Copy)Document2 pages03 Meninges, Neuron, NMJ, CSF (Students Copy)Hillary DangananNo ratings yet
- Color Atlas of BrainDocument8 pagesColor Atlas of BrainMWNo ratings yet
- Frontal LobeDocument39 pagesFrontal LobeVIKAS PUNIANo ratings yet
- Open AnatomyDocument50 pagesOpen AnatomyalukiasuNo ratings yet
- Ilovepdf MergedDocument65 pagesIlovepdf Mergedrogegir407No ratings yet
- 17 Skull Mandible Vertebrae DR - GosaiDocument69 pages17 Skull Mandible Vertebrae DR - GosaiDr.B.B.GosaiNo ratings yet
- Deiuliis2019 VERTEBRADOS LA PERCA 4Document10 pagesDeiuliis2019 VERTEBRADOS LA PERCA 4Fernando Melo GarciaNo ratings yet
- L15 CerebrumDocument27 pagesL15 CerebrumEvilneko1No ratings yet
- Mini AtlasDocument28 pagesMini AtlasAlex WoodwardNo ratings yet
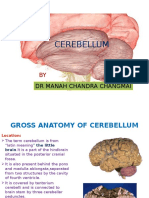
- Cerebellum: DR Manah Chandra ChangmaiDocument60 pagesCerebellum: DR Manah Chandra Changmainairda12No ratings yet
- 3 Cross-Sectional Anatomy With Corresponding CT Images: 3.1 Head, Brain, and Visceral CraniumDocument2 pages3 Cross-Sectional Anatomy With Corresponding CT Images: 3.1 Head, Brain, and Visceral CraniumSabina FarhinNo ratings yet
- Elbow Complex (Kinesiology)Document2 pagesElbow Complex (Kinesiology)Kimmybee Garcia50% (2)
- Nerve - H&N PracticeDocument7 pagesNerve - H&N PracticeThục MỹNo ratings yet
- 02 SkullDocument19 pages02 Skullapi-19641337No ratings yet
- Anatomia para Estética Parte 1Document45 pagesAnatomia para Estética Parte 1Rodolpho VilelaNo ratings yet
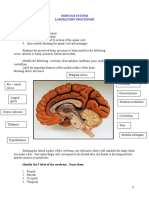
- Materials Needed:: Nervous System Laboratory ProcedureDocument7 pagesMaterials Needed:: Nervous System Laboratory ProcedureRegina GambayanNo ratings yet
- Chapter 5Document41 pagesChapter 5Reneilwe MoshidiNo ratings yet
- Book 1 - Sketch MedDocument220 pagesBook 1 - Sketch MedScribdTranslationsNo ratings yet
- 1 - The SkullDocument68 pages1 - The Skullaboody omerNo ratings yet
- p6. The Cerebral CortexDocument99 pagesp6. The Cerebral CortexGedefaw TegegneNo ratings yet
- Lab+Exam+1+Worksheet+ +terminology 1Document11 pagesLab+Exam+1+Worksheet+ +terminology 1jharvillgreeneNo ratings yet
- Cerebral Cortex1Document37 pagesCerebral Cortex1xwrmaliaramNo ratings yet
- (849-853) Component PartsDocument5 pages(849-853) Component PartsMỉm CườiNo ratings yet
- Poster01 Skeletal-System-Anterior Ledger11x17Document1 pagePoster01 Skeletal-System-Anterior Ledger11x17jeffrin alexanderNo ratings yet
- Gross Structure of The BrainDocument60 pagesGross Structure of The BrainPurnomo Ponco NugrohoNo ratings yet
- Head & Neck ManualDocument176 pagesHead & Neck Manualdrfizaaliblogs632No ratings yet
- Anatomy of The Human BodyDocument6 pagesAnatomy of The Human BodyYara AmadeNo ratings yet
- HAPMLSDocument4 pagesHAPMLSKrea Shin MatsuokaNo ratings yet
- Scalp (2) PP - 240218 - 203325Document16 pagesScalp (2) PP - 240218 - 203325Hager AbouzaidNo ratings yet
- 3 Face and Parotid SlidesDocument23 pages3 Face and Parotid SlidesAmbg GhalyNo ratings yet
- معمل upper limb د علي البغدادي الجزء الاولDocument8 pagesمعمل upper limb د علي البغدادي الجزء الاولAbdulaziz MohammedNo ratings yet
- Osteology Summary NotesDocument25 pagesOsteology Summary Notesmilex75862No ratings yet
- Bones & Joints of Head & NeckDocument91 pagesBones & Joints of Head & NeckSalman KhanNo ratings yet
- Lecture 12 Osteology Upper LimbDocument25 pagesLecture 12 Osteology Upper LimbaidfajdfiajNo ratings yet
- Lab 4 - ForearmDocument44 pagesLab 4 - ForearmTayai LengimaNo ratings yet
- The Axial SkeletonDocument57 pagesThe Axial SkeletonJudithNo ratings yet
- Cerebral Cortex: Medical Neuroscience Dr. WiegandDocument24 pagesCerebral Cortex: Medical Neuroscience Dr. WiegandUzma KhanNo ratings yet
- CephalometricsDocument141 pagesCephalometricsshyama pramodNo ratings yet
- Chapter 11-Human Body SystemDocument48 pagesChapter 11-Human Body SystemTrần MạnhNo ratings yet
- Transportation in Animals and Plants Worksheet 8Document2 pagesTransportation in Animals and Plants Worksheet 8Parul ShahNo ratings yet
- Endodontic Orthodontic Relationships A Review of Integrated Treatment PlanningDocument18 pagesEndodontic Orthodontic Relationships A Review of Integrated Treatment PlanningAlfonso SernaNo ratings yet
- Physics of The Eye and VisionDocument3 pagesPhysics of The Eye and VisionhusseinNo ratings yet
- Physical Therapy Neuroscience Central Somatosensory Lecture POSTDocument43 pagesPhysical Therapy Neuroscience Central Somatosensory Lecture POSTashlyn granthamNo ratings yet
- 1678816971346634Document47 pages1678816971346634SophiyaNo ratings yet
- Explain Why The Larger Waves Seen On The Oscilloscope Represent The Ventricular ContractionDocument1 pageExplain Why The Larger Waves Seen On The Oscilloscope Represent The Ventricular ContractionFatimah Shellya ShahabNo ratings yet
- Jacqueline Levana Huliselan (X511)Document11 pagesJacqueline Levana Huliselan (X511)Jacqueline Levana HuliselanNo ratings yet
- 2.0 Topic 2 Epithelium Non-Epithelial Cells (HDD30203)Document41 pages2.0 Topic 2 Epithelium Non-Epithelial Cells (HDD30203)AugustNo ratings yet
- Week 4 - Plant Structure and FunctionsDocument50 pagesWeek 4 - Plant Structure and FunctionsPrincess De LeonNo ratings yet
- Anatomy Physiology of The BrainDocument107 pagesAnatomy Physiology of The BrainRadha RamineniNo ratings yet
- CELL CITY PROJECT - You Are The Architect: Manuel S. Enverga University Foundation Candelaria, IncDocument3 pagesCELL CITY PROJECT - You Are The Architect: Manuel S. Enverga University Foundation Candelaria, IncDEL ROSARIO GWEN ASHLEY R.No ratings yet
- Plant Responses and GrowthDocument3 pagesPlant Responses and GrowthJung Ahn HongNo ratings yet
- Anatomy and Physiology 6th Edition Gunstream Test Bank Full Chapter PDFDocument42 pagesAnatomy and Physiology 6th Edition Gunstream Test Bank Full Chapter PDFserenafinnodx100% (13)
- CHAPTER 17 NutritionDocument4 pagesCHAPTER 17 Nutritionharry potingNo ratings yet
- Ged Life Science 2Document27 pagesGed Life Science 2mallikammu12No ratings yet
- Smooth Muscle Physiology by Bushra Akhtar Dhillon (07.04.2020)Document24 pagesSmooth Muscle Physiology by Bushra Akhtar Dhillon (07.04.2020)Laraib SohailNo ratings yet
- Psychology of StressDocument39 pagesPsychology of StressMikhaeel RizkNo ratings yet
- 54BS482 CarterDocument2 pages54BS482 CartermarlenyNo ratings yet
- The Enthesis: A Review of The Tendon-To-Bone InsertionDocument10 pagesThe Enthesis: A Review of The Tendon-To-Bone InsertionLuis Briones Lau.LiNo ratings yet
- Drug InteractionDocument18 pagesDrug InteractionBp hoNo ratings yet
- Kami Export - Aubrianna Labonte-Postell - CellStructure GizmoDocument5 pagesKami Export - Aubrianna Labonte-Postell - CellStructure Gizmoaubrianna.postellNo ratings yet
- ALLERGIESDocument5 pagesALLERGIESKebbewar KhaledNo ratings yet
- Pixl Knowit!: Gcse BiologyDocument85 pagesPixl Knowit!: Gcse BiologyKausar AhmedNo ratings yet
- K Devi Sankar, Assistant Professor of Anatomy, Narayana Medical College, Nellore, Andhra Pradesh, IndiaDocument35 pagesK Devi Sankar, Assistant Professor of Anatomy, Narayana Medical College, Nellore, Andhra Pradesh, IndialesansharNo ratings yet
- Legal Medicine Quiz SolisDocument1 pageLegal Medicine Quiz SolisMartin Clyde PaglinawanNo ratings yet
- Super Simplified Pathology Hematology - Dr. Priyanka SachdevDocument500 pagesSuper Simplified Pathology Hematology - Dr. Priyanka SachdevMohd SaquibNo ratings yet
- Microbial Ecology and Diversity AssignmentDocument19 pagesMicrobial Ecology and Diversity AssignmentGutu EmiruNo ratings yet
- Frontiers - Rheumatic Heart Valve Disease Pathophysiology and Underlying MechanismsDocument33 pagesFrontiers - Rheumatic Heart Valve Disease Pathophysiology and Underlying MechanismsJAHNELLA XYRINE PERITONo ratings yet
- Introduction To Osteology - 21-04-2022Document29 pagesIntroduction To Osteology - 21-04-2022ANN MARIYA MONCYNo ratings yet