Download as pdf or txt
You might also like
- A Level Biology For OCR A (Double Paged)Document360 pagesA Level Biology For OCR A (Double Paged)squishyboitae33% (3)
- EEB 162 Final Study GuideDocument13 pagesEEB 162 Final Study GuideSydney ChangNo ratings yet
- Plant PhysiologyDocument95 pagesPlant PhysiologyAlvin B BuenconsejoNo ratings yet
- Phloem - WikipediaDocument27 pagesPhloem - WikipediaBashiir NuurNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument20 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For Scientistsasmeraamde21No ratings yet
- Plant Cells Vs Animal CellsDocument51 pagesPlant Cells Vs Animal CellsDivyakumar PatelNo ratings yet
- Content: - IntroductionDocument12 pagesContent: - IntroductionSaziya QuadriNo ratings yet
- Cell and TissuesDocument13 pagesCell and TissuesJeffrish Doni EzhilNo ratings yet
- Biology SechayaDocument164 pagesBiology SechayaJulien KhalilNo ratings yet
- Phloem: - Sieve Tube Elements (Angiosperms) - Sieve Cells (Gymnosperms)Document9 pagesPhloem: - Sieve Tube Elements (Angiosperms) - Sieve Cells (Gymnosperms)xbNo ratings yet
- Fundamentals of Crop PhysiologyDocument19 pagesFundamentals of Crop PhysiologyRsNo ratings yet
- 9.2 Biology HL PresentationDocument21 pages9.2 Biology HL PresentationnanaNo ratings yet
- Experiment 1 Cell BioDocument7 pagesExperiment 1 Cell BioWinserng 永森No ratings yet
- Structure and Functions of Plant TissuesDocument8 pagesStructure and Functions of Plant Tissuesdina aribahNo ratings yet
- Plastid: Plastids in PlantsDocument9 pagesPlastid: Plastids in Plantsrs6622No ratings yet
- SF - MD.0145.T.22 Ally Omary MwambelaDocument10 pagesSF - MD.0145.T.22 Ally Omary MwambelaAlly OmaryNo ratings yet
- FT1043 Lecture 1Document40 pagesFT1043 Lecture 1Norzhafarina M. IsmailNo ratings yet
- UP Diliman - Geol 130 Plants ReportDocument94 pagesUP Diliman - Geol 130 Plants ReportMac ServitoNo ratings yet
- Lec1 PlantCellDocument30 pagesLec1 PlantCellNaHuynJungNo ratings yet
- Lec1 PlantCellDocument30 pagesLec1 PlantCellMinh ChâuNo ratings yet
- Cell OrganellesDocument7 pagesCell OrganellesAdit KumarNo ratings yet
- The Following Is A Glossary of Plant Cell Anatomy Terms.: Plant Cells Cristae Mitochondria ChloroplastsDocument6 pagesThe Following Is A Glossary of Plant Cell Anatomy Terms.: Plant Cells Cristae Mitochondria ChloroplastsHitesh MoreNo ratings yet
- Chapter 4: Cell Membranes and Transport: SummaryDocument4 pagesChapter 4: Cell Membranes and Transport: SummaryMerimaNo ratings yet
- Plant Organology Notes PDFDocument18 pagesPlant Organology Notes PDFCaitlin SnymanNo ratings yet
- Protozoa: Characteristics, Reproduction and ClassificationDocument13 pagesProtozoa: Characteristics, Reproduction and ClassificationAristeo EbioNo ratings yet
- Chloroplasts: Microsoft ® Encarta ® 2009. © 1993-2008 Microsoft Corporation. All Rights ReservedDocument14 pagesChloroplasts: Microsoft ® Encarta ® 2009. © 1993-2008 Microsoft Corporation. All Rights ReservedMelissa R.BartolayNo ratings yet
- Cell Organelles JigsawDocument10 pagesCell Organelles JigsawSusan VictoriaNo ratings yet
- Cells 3Document66 pagesCells 3KEANNA RUBIANo ratings yet
- Topic 4 Edexcel - 240101 - 190205Document57 pagesTopic 4 Edexcel - 240101 - 190205Ovina RanasingheNo ratings yet
- Plant Transport Paper 2Document2 pagesPlant Transport Paper 2Heneral Drew de LunaNo ratings yet
- Chloroplast NotesDocument4 pagesChloroplast NotesAnanda100% (1)
- Plant Cell - WikipediaDocument33 pagesPlant Cell - WikipediaNaniNo ratings yet
- Biology Nmat New SetDocument8 pagesBiology Nmat New SetOrlan Defensor Balano88% (16)
- Plant Cell - WikipediaDocument32 pagesPlant Cell - WikipediaSherelyn Bernardo100% (1)
- Biology For Prelims (Sscstudy - Com)Document385 pagesBiology For Prelims (Sscstudy - Com)Abhisar KhokharNo ratings yet
- Meristem Parenchyma Sclerenchyma 5: Figure 5: Cell Types and TissuesDocument8 pagesMeristem Parenchyma Sclerenchyma 5: Figure 5: Cell Types and Tissuessandy waskitoNo ratings yet
- TitleDocument10 pagesTitleWei Loon100% (1)
- Vascular Tissue in PlantsDocument30 pagesVascular Tissue in Plantsdinda0% (1)
- Unit 3. Cellular Basis of Life-2Document13 pagesUnit 3. Cellular Basis of Life-2Kerala MekuriyaNo ratings yet
- Cell - BauyonDocument7 pagesCell - BauyonbauyonanchieNo ratings yet
- BSEDSCI 13 (Algea) Patinga, Christine Mae V.Document3 pagesBSEDSCI 13 (Algea) Patinga, Christine Mae V.Christian PatingaNo ratings yet
- Mitochondria: JWBK011-03 JWBK011-Hogg August 12, 2005 15:52 Char Count 0 Cell Structure and OrganisationDocument2 pagesMitochondria: JWBK011-03 JWBK011-Hogg August 12, 2005 15:52 Char Count 0 Cell Structure and OrganisationJona KoçiNo ratings yet
- Cell Unit of LifeDocument8 pagesCell Unit of LifeNavaneeth KrishnanNo ratings yet
- Lection 1 and 2Document2 pagesLection 1 and 2MariaNo ratings yet
- Bio2 4TH QTRDocument11 pagesBio2 4TH QTRjames DeeNo ratings yet
- Prerequisite Knowledge of The Structure of The Plant.: PlantsDocument11 pagesPrerequisite Knowledge of The Structure of The Plant.: PlantsJohn Bryan JamisonNo ratings yet
- Extra SCDocument2 pagesExtra SCuwj97No ratings yet
- Cell Class Viii - 3 CompetitiveDocument23 pagesCell Class Viii - 3 Competitiverajesh duaNo ratings yet
- Transport of Materials in Flowering PlantsDocument22 pagesTransport of Materials in Flowering PlantsMd SafwatNo ratings yet
- Lab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeDocument4 pagesLab 4 Drawing of A Cross Section of A Ligustrum Plant Cell Under A Light MicroscopeMichael TimsonNo ratings yet
- Document 4Document11 pagesDocument 4pragany.saiNo ratings yet
- Cell Organeeles IDocument7 pagesCell Organeeles ISaumya GopalNo ratings yet
- The Plasma (Cytoplasmic) MembraneDocument6 pagesThe Plasma (Cytoplasmic) Membraneprism1702No ratings yet
- Class 11 Biology Chapter 8 - Revision NotesDocument10 pagesClass 11 Biology Chapter 8 - Revision NotesHARKIRIT KAUR100% (1)
- Chloroplast 1.2Document5 pagesChloroplast 1.2Ishita Kumari100% (1)
- Bioloy Notes From NCERT (Mahendra Coaching NotesDocument385 pagesBioloy Notes From NCERT (Mahendra Coaching NotesBabu VermaNo ratings yet
- Lab 1 Plant MorphologyDocument5 pagesLab 1 Plant MorphologyNabilah SyahirahNo ratings yet
- Plant Cell StructureDocument3 pagesPlant Cell Structurengozi duffrinNo ratings yet
- How Plants Grow - With Information on the Biology of Plant Cells, Roots, Leaves and FlowersFrom EverandHow Plants Grow - With Information on the Biology of Plant Cells, Roots, Leaves and FlowersNo ratings yet
- Morfologi Bunga Dan Viabilitas Serbuk Sari Berbagai Aksesi PAMELO (Citrus Maxima (Burm.) Merr.)Document9 pagesMorfologi Bunga Dan Viabilitas Serbuk Sari Berbagai Aksesi PAMELO (Citrus Maxima (Burm.) Merr.)Tsamara Khansa AzzahraNo ratings yet
- KDPTG 2020Document33 pagesKDPTG 2020AnonimNo ratings yet
- Grade 5 Cot Lesson PlanDocument6 pagesGrade 5 Cot Lesson PlanSAMUEL SISON ZONIO100% (1)
- LeavesDocument34 pagesLeavesGuirelaNo ratings yet
- Fruit ClassificationDocument1 pageFruit ClassificationMuhd Azhar ZuhairyNo ratings yet
- AngiospermDocument5 pagesAngiospermAbdul BasitNo ratings yet
- Lecture 13Document8 pagesLecture 13hotlipz_126No ratings yet
- Morphological and Anatomical Characteristic of Pisang Awak (Musa Paradisiaca Cv. Awak) in West KalimantanDocument7 pagesMorphological and Anatomical Characteristic of Pisang Awak (Musa Paradisiaca Cv. Awak) in West KalimantanMaria AnggieNo ratings yet
- Worksheet Science Kelas 5 Ke 3Document4 pagesWorksheet Science Kelas 5 Ke 3Arif Prasetyo WibowoNo ratings yet
- Transportation in PlantsDocument8 pagesTransportation in PlantsRoseNo ratings yet
- 20170918140947SBC 3023 - TBB 2033beditednew4Document36 pages20170918140947SBC 3023 - TBB 2033beditednew4Leena MuniandyNo ratings yet
- Lab Rep 4Document27 pagesLab Rep 4Reign RosadaNo ratings yet
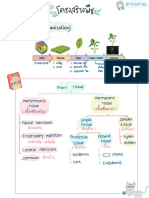
- สรุปโครงสร้างพืชDocument4 pagesสรุปโครงสร้างพืชรินบิNo ratings yet
- Anatomi Dan Morfologi Tumbuhan (Plant Anatomy and Morphology) BY: Dr. Nor Nafizah Mohd NoorDocument53 pagesAnatomi Dan Morfologi Tumbuhan (Plant Anatomy and Morphology) BY: Dr. Nor Nafizah Mohd NoorLeena MuniandyNo ratings yet
- Botany Plant Anatomy SEM II H SEM I GDocument34 pagesBotany Plant Anatomy SEM II H SEM I GMartha-Angella MassaquoiNo ratings yet
- Morphological and Anatomical Characters of Piper Betle L. From Madaya Township, Mandalay RegionDocument19 pagesMorphological and Anatomical Characters of Piper Betle L. From Madaya Township, Mandalay RegionAxel PangloliNo ratings yet
- DownloadDocument10 pagesDownloadKatrin Choisy D. PascualNo ratings yet
- Studies in Neotropical Vochysiaceae: A New Species of Vochysia (Section Ciliantha) From A Pluvial Forest of Western ColombiaDocument13 pagesStudies in Neotropical Vochysiaceae: A New Species of Vochysia (Section Ciliantha) From A Pluvial Forest of Western ColombiabioandreyNo ratings yet
- XI CH - 06 Anatomy of Flowering Plants - 220605 - 233239Document17 pagesXI CH - 06 Anatomy of Flowering Plants - 220605 - 233239Annu BaijalNo ratings yet
- FlowersDocument10 pagesFlowersAmina Zeghar BNo ratings yet
- Janka Hardness Test: Navigation SearchDocument5 pagesJanka Hardness Test: Navigation SearchSafy SerenityNo ratings yet
- 307 48 ET V1 S1 - File1 PDFDocument22 pages307 48 ET V1 S1 - File1 PDFaaaatniNo ratings yet
- Pedoman Kerja Proses Produksi Kayu Lapis BagusDocument25 pagesPedoman Kerja Proses Produksi Kayu Lapis BagusliaNo ratings yet
- Structure of Ovule and TypesDocument10 pagesStructure of Ovule and TypesJemson KhundongbamNo ratings yet
- Pollination Project New 2Document30 pagesPollination Project New 2SagarNo ratings yet
- TamarindDocument31 pagesTamarind20UHR011 Mohan KumarNo ratings yet
- Aestivation & Inflorascence ....Document13 pagesAestivation & Inflorascence ....Abhimanyu PandeyNo ratings yet
- Science Grade 3 ExamDocument2 pagesScience Grade 3 ExamAlaNo ratings yet
- Entry Bench Cutting List: QT. T W LDocument7 pagesEntry Bench Cutting List: QT. T W Ldony ramdhaniNo ratings yet
- Sexual Reproduction in Flowering PlantsDocument66 pagesSexual Reproduction in Flowering Plantsshobitgup100% (1)