Download as pdf or txt
You might also like
- Gpat Opat Omae Microbio COMPILATIONDocument136 pagesGpat Opat Omae Microbio COMPILATIONDENTAL REVIEWER ONLYNo ratings yet
- NBPME Practice Tests 2005Document39 pagesNBPME Practice Tests 2005Pat TNo ratings yet
- Biogas Plant: Biogas Digester Design, Gas Production and PurificationFrom EverandBiogas Plant: Biogas Digester Design, Gas Production and PurificationRating: 3 out of 5 stars3/5 (1)
- Schematic Diagram On The 4 Types of ShockDocument5 pagesSchematic Diagram On The 4 Types of Shockgodgiven25100% (5)
- 951-Article Text-2783-1-10-20221231Document6 pages951-Article Text-2783-1-10-20221231MohamedTaherNo ratings yet
- 5 - Biodiesel, Biogas and Fermentable Sugars Production From Spent Coffee Grounds - A Cascade Biorefinery ApproachDocument8 pages5 - Biodiesel, Biogas and Fermentable Sugars Production From Spent Coffee Grounds - A Cascade Biorefinery Approachjackeline15No ratings yet
- Integrating Sugarcane Molasses Into Sequential Cellulosic Biofuel Production Based On SSF Process of High Solid LoadingDocument9 pagesIntegrating Sugarcane Molasses Into Sequential Cellulosic Biofuel Production Based On SSF Process of High Solid LoadingSamuel GamorNo ratings yet
- Delgado-Noboa_et_al_2021 (1)Document7 pagesDelgado-Noboa_et_al_2021 (1)ALONDRA NICOLE GUZMAN FUENTESNo ratings yet
- Jof 08 00395 With CoverDocument23 pagesJof 08 00395 With CoverVitor ArsenioNo ratings yet
- FermentationDocument18 pagesFermentationCrissy DionesNo ratings yet
- Jof 08 00395 v2Document22 pagesJof 08 00395 v2IslamProNo ratings yet
- A Waldron ArticleDocument12 pagesA Waldron ArticleThịnh Vương KiếnNo ratings yet
- Bio-Ethanol ProductionDocument11 pagesBio-Ethanol ProductionRavi ShankarNo ratings yet
- 1 s2.0 S2589014X21003133 MainDocument13 pages1 s2.0 S2589014X21003133 MainFelipe RojasNo ratings yet
- Bioresource TechnologyDocument7 pagesBioresource TechnologyZAk1No ratings yet
- Pretreatment and Hydrolysis of Cellulosic Agricultural Wastes With A Cellulase-Producing Streptomyces For Bioethanol ProductionDocument7 pagesPretreatment and Hydrolysis of Cellulosic Agricultural Wastes With A Cellulase-Producing Streptomyces For Bioethanol ProductionAndré Felipe SilvaNo ratings yet
- 1 s2.0 S094450132030402X MainDocument16 pages1 s2.0 S094450132030402X Mainbiovijay101No ratings yet
- Energy: Wei Luo, Zhangmin Zhao, Hepeng Pan, Lankun Zhao, Chuangao Xu, Xiaobin YuDocument9 pagesEnergy: Wei Luo, Zhangmin Zhao, Hepeng Pan, Lankun Zhao, Chuangao Xu, Xiaobin YuCarlos Ruben Suarez GavidiaNo ratings yet
- Effectiveness of Low-Cost Bioreactors Integrated With A Gas Stripping System For Butanol Fermentation From Sugarcane Molasses by Clostridium BeijerinckiiDocument13 pagesEffectiveness of Low-Cost Bioreactors Integrated With A Gas Stripping System For Butanol Fermentation From Sugarcane Molasses by Clostridium BeijerinckiiMarianValdiviaNo ratings yet
- 1 s2.0 S2215017X23000358 MainDocument11 pages1 s2.0 S2215017X23000358 Mainsrajasree98No ratings yet
- Artigo 1Document7 pagesArtigo 1Otavio Lucas Alves da SilvaNo ratings yet
- sALAKAM23 SUPERDocument9 pagessALAKAM23 SUPERYoel Alfonso AcostaNo ratings yet
- A Novel Cell Factory For Efficient Production of Ethanol From Dairy WasteDocument11 pagesA Novel Cell Factory For Efficient Production of Ethanol From Dairy WasteVitor ArsenioNo ratings yet
- Recuperación y Caracterización de Etanol Celulósico A Partir de La Fermentación de Bagazo de Caña de AzúcarDocument9 pagesRecuperación y Caracterización de Etanol Celulósico A Partir de La Fermentación de Bagazo de Caña de AzúcarFabian VelasquezNo ratings yet
- COZMA AdrianaDocument7 pagesCOZMA Adrianakeepemdown17No ratings yet
- A Review of Bioethanol Production From Plant-Based Waste Biomass by Yeast FermentationDocument15 pagesA Review of Bioethanol Production From Plant-Based Waste Biomass by Yeast FermentationMi PensNo ratings yet
- Bioethanol Production From Renewable Raw Materials and Its Separation and Purification - A ReviewDocument38 pagesBioethanol Production From Renewable Raw Materials and Its Separation and Purification - A Reviewnabeelkhaliq323No ratings yet
- Fruit Leather Inggris 5 DoneDocument3 pagesFruit Leather Inggris 5 DonefitriNo ratings yet
- Biochemical Engineering JournalDocument13 pagesBiochemical Engineering JournalolaragoncNo ratings yet
- Production of Lactic Acid On Liquid Distillery Stillage Proizvodnja Mlečne Kiseline Na Tečnoj Destilerijskoj DžibriDocument5 pagesProduction of Lactic Acid On Liquid Distillery Stillage Proizvodnja Mlečne Kiseline Na Tečnoj Destilerijskoj DžibriMathilda PasaribuNo ratings yet
- 2013 Scottetal Biotechnol Biofuels 6171Document17 pages2013 Scottetal Biotechnol Biofuels 6171Martha AlbaNo ratings yet
- 2.bio Butanol ProductionDocument11 pages2.bio Butanol Productionsupercubos1No ratings yet
- Producao Entanol ResiduosleiteDocument10 pagesProducao Entanol ResiduosleiteVitor ArsenioNo ratings yet
- The Production of Fuel-Grade Bioethanol From Cassava Starch: A Conceptual Process DesignDocument13 pagesThe Production of Fuel-Grade Bioethanol From Cassava Starch: A Conceptual Process DesignPutu Yoga100% (1)
- 10 1016@j Rser 2017 01 151 PDFDocument12 pages10 1016@j Rser 2017 01 151 PDFmochdimaskuNo ratings yet
- A New Method For Industrial Production of 2,3-Butanediol: Lan Ge, Xiaomin Wu, Jianwen Chen, Jialin WuDocument3 pagesA New Method For Industrial Production of 2,3-Butanediol: Lan Ge, Xiaomin Wu, Jianwen Chen, Jialin WuChristopher BalalembangNo ratings yet
- Bioethanol Production FromDocument12 pagesBioethanol Production FromLeta DerejeNo ratings yet
- Renewable Energy: Naseeha A. Chohan, G.S. Aruwajoye, Y. Sewsynker-Sukai, E.B. Gueguim KanaDocument10 pagesRenewable Energy: Naseeha A. Chohan, G.S. Aruwajoye, Y. Sewsynker-Sukai, E.B. Gueguim KanaAyush SinghNo ratings yet
- Biore Source Technology: Shih-Hsin Ho, Shu-Wen Huang, Chun-Yen Chen, Tomohisa Hasunuma, Akihiko Kondo, Jo-Shu ChangDocument8 pagesBiore Source Technology: Shih-Hsin Ho, Shu-Wen Huang, Chun-Yen Chen, Tomohisa Hasunuma, Akihiko Kondo, Jo-Shu ChangViona WidyaNo ratings yet
- Cassava Pulp Enzymatic Hydrolysis Process As A Preliminary Step in Bio-AlcoholsDocument7 pagesCassava Pulp Enzymatic Hydrolysis Process As A Preliminary Step in Bio-AlcoholsProfessor Douglas TorresNo ratings yet
- Bioresource Technology: Yanan Zhang, Tristan R. Brown, Guiping Hu, Robert C. BrownDocument8 pagesBioresource Technology: Yanan Zhang, Tristan R. Brown, Guiping Hu, Robert C. BrownAnonymous sVheFhq9PNo ratings yet
- Development of A Novel, Robust and Cost-Efficient Process For Valorizing Dairy Waste Exemplified by Ethanol ProductionDocument10 pagesDevelopment of A Novel, Robust and Cost-Efficient Process For Valorizing Dairy Waste Exemplified by Ethanol ProductionwfkamNo ratings yet
- 9 A Biorefinery ApproachDocument15 pages9 A Biorefinery ApproachJavier Andres Davila RinconNo ratings yet
- Mekanism Antibakteri 3Document15 pagesMekanism Antibakteri 3Sintha HapsariNo ratings yet
- Chemosphere: Cerevisiae and Beet Molasses Inoculum A Biomass Based InvestigationDocument6 pagesChemosphere: Cerevisiae and Beet Molasses Inoculum A Biomass Based InvestigationDika SetiaNo ratings yet
- Energies: Second Generation Ethanol Production From Brewers' Spent GrainDocument12 pagesEnergies: Second Generation Ethanol Production From Brewers' Spent GrainmulerNo ratings yet
- BiobutanolDocument11 pagesBiobutanolPabloDoriaNo ratings yet
- A Simple Scaled Down System To Mimic The Industrial Production of First Generation Fuel Ethanol in BrazilDocument13 pagesA Simple Scaled Down System To Mimic The Industrial Production of First Generation Fuel Ethanol in BrazilTamiresFariaNo ratings yet
- Intensive Production of Carboxylic Acids Using C. Butyricum in A Membrane Bioreactor (MBR)Document24 pagesIntensive Production of Carboxylic Acids Using C. Butyricum in A Membrane Bioreactor (MBR)Threse AreolaNo ratings yet
- Tutorial 2Document3 pagesTutorial 2Lê PhươngNo ratings yet
- Bioresource Technology: Raveendran Sindhu, Parameswaran Binod, Ashok PandeyDocument7 pagesBioresource Technology: Raveendran Sindhu, Parameswaran Binod, Ashok PandeyMuhammad Fakhrizal Fahmi100% (1)
- 1 s2.0 S0959652622029262 MainDocument9 pages1 s2.0 S0959652622029262 MainBernard SNo ratings yet
- Bioethanol Sugarcane Vs Sugarcane BagasseDocument10 pagesBioethanol Sugarcane Vs Sugarcane Bagassesunisht KumarNo ratings yet
- Bioethanol From Sweet Potato Clones at Pruning Age and Yeast ConcentrationsDocument7 pagesBioethanol From Sweet Potato Clones at Pruning Age and Yeast ConcentrationsMamta AgarwalNo ratings yet
- Bioethanol Production From Corn Stover Using Saccharomyces CerevisiaeDocument5 pagesBioethanol Production From Corn Stover Using Saccharomyces CerevisiaeNikhil wadicharNo ratings yet
- Techno-Economic Evaluation of 2 Generation Bioethanol Production From Sugar Cane Bagasse and Leaves Integrated With The Sugar-Based Ethanol ProcessDocument18 pagesTechno-Economic Evaluation of 2 Generation Bioethanol Production From Sugar Cane Bagasse and Leaves Integrated With The Sugar-Based Ethanol ProcessPanda FozolNo ratings yet
- Dave2019 PDFDocument14 pagesDave2019 PDFuchechukwu ikebuduNo ratings yet
- Lactosuero 5 NoDocument14 pagesLactosuero 5 NoMarco Antonio Salcedo HinojosaNo ratings yet
- Bioethanol Production From Scenedesmus Obliquus SugarsDocument11 pagesBioethanol Production From Scenedesmus Obliquus SugarsLAURA LUC�A ATENCIA CASTILLONo ratings yet
- Assignment 1 Che 544 - Group 6Document29 pagesAssignment 1 Che 544 - Group 6atikah hasnor100% (1)
- Fenrg 11 1183035Document20 pagesFenrg 11 1183035Vitor ArsenioNo ratings yet
- Advances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsFrom EverandAdvances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsLalit Kumar SinghNo ratings yet
- Advances in Biofeedstocks and Biofuels, Volume 1: Biofeedstocks and Their ProcessingFrom EverandAdvances in Biofeedstocks and Biofuels, Volume 1: Biofeedstocks and Their ProcessingLalit Kumar SinghNo ratings yet
- Excerpt From Spark - The Revolutionary Science of Exercise and The Brain LoyolaDocument5 pagesExcerpt From Spark - The Revolutionary Science of Exercise and The Brain LoyolaValerio Ernesto CòlibroNo ratings yet
- Lee 2010Document9 pagesLee 2010Hoàng NhậtNo ratings yet
- Pharmacology ReviewersDocument5 pagesPharmacology ReviewersNeisha Halil VillarealNo ratings yet
- Opportunities For Early Intervention Based On Theory, Basic Neuroscience, and Clinical ScienceDocument13 pagesOpportunities For Early Intervention Based On Theory, Basic Neuroscience, and Clinical ScienceRama BayuNo ratings yet
- Biology PracticalDocument21 pagesBiology Practicaldhruvps818No ratings yet
- Document On Flowering of PlantDocument36 pagesDocument On Flowering of PlantIHB BHARTINo ratings yet
- Ha NotesDocument34 pagesHa NotesLyssa KateNo ratings yet
- Breathing Process PDFDocument3 pagesBreathing Process PDFDianeNo ratings yet
- The Intestinal Microbiota Fuelling Metabolic Inflammation: Herbert Tilg, Niv Zmora, Timon E. Adolph and Eran ElinavDocument15 pagesThe Intestinal Microbiota Fuelling Metabolic Inflammation: Herbert Tilg, Niv Zmora, Timon E. Adolph and Eran ElinavAndrés Felipe Zapata MurielNo ratings yet
- Biomechanics of NervesDocument65 pagesBiomechanics of NervesZinneRah RahManNo ratings yet
- Скелетна ммускулна клетка - устройство на саркомерDocument4 pagesСкелетна ммускулна клетка - устройство на саркомерMartin PetkovNo ratings yet
- The Neuron: Vce Psychology - Unit 3 Aos 1Document22 pagesThe Neuron: Vce Psychology - Unit 3 Aos 1C0SMICALCHEMISTNo ratings yet
- Second Quarter ExamDocument7 pagesSecond Quarter ExamJelyn RamirezNo ratings yet
- Class Pterobranchia (Sea Angels) Class Enteropneusta (Acorn Worms)Document9 pagesClass Pterobranchia (Sea Angels) Class Enteropneusta (Acorn Worms)muhammadismailNo ratings yet
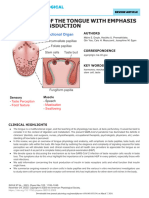
- Physiology of The Tongue With Emphasis On Taste TransductionDocument55 pagesPhysiology of The Tongue With Emphasis On Taste Transductionfajriatinazula02No ratings yet
- Biophysical ProfileDocument8 pagesBiophysical ProfileAmritaNo ratings yet
- Analizador de Bioquímica Automatizado BiobaseDocument7 pagesAnalizador de Bioquímica Automatizado BiobaseGenesis DiazNo ratings yet
- Anemia OsamaDocument57 pagesAnemia Osamaosamafoud7710No ratings yet
- Connie Mosuela MS FE 1Document3 pagesConnie Mosuela MS FE 1Tricia Jane OpinaldoNo ratings yet
- Anatomy and Physiology GMCDocument15 pagesAnatomy and Physiology GMCsamantha tanoNo ratings yet
- Science 6 q2 Mod1Document20 pagesScience 6 q2 Mod13tj internetNo ratings yet
- Passive Transport - Virtual LabDocument27 pagesPassive Transport - Virtual LabHalleli CastilloNo ratings yet
- Physiology Final Exam - Glory 2017 PDFDocument14 pagesPhysiology Final Exam - Glory 2017 PDFMohammad BarakatNo ratings yet
- Unit 305 AssignmentDocument9 pagesUnit 305 Assignmenthiba.khaliddd0No ratings yet
- Neuroanatomy Mcqs by Drs of 2027-28Document19 pagesNeuroanatomy Mcqs by Drs of 2027-28muhammadshayan416No ratings yet
- November 2021 (v3) QP - Paper 2 CAIE Biology IGCSEDocument16 pagesNovember 2021 (v3) QP - Paper 2 CAIE Biology IGCSEanahmadNo ratings yet
- Lu III 0910 2ndsem Tentative SchedDocument30 pagesLu III 0910 2ndsem Tentative Schedaa628No ratings yet