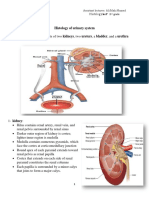
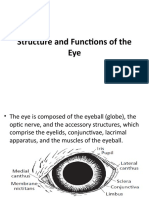
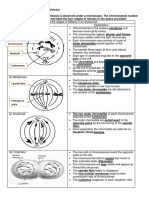
Download as pdf or txt
You might also like
- Preclinical Anatomy Review 2023: For USMLE Step 1 and COMLEX-USA Level 1From EverandPreclinical Anatomy Review 2023: For USMLE Step 1 and COMLEX-USA Level 1Rating: 5 out of 5 stars5/5 (2)
- WwwmensruationDocument12 pagesWwwmensruationTamar GrdzelishviliNo ratings yet
- 1 s2.0 S0093691X22003934 MainDocument8 pages1 s2.0 S0093691X22003934 MainMarco AsprónNo ratings yet
- Regeneration of The LiverDocument29 pagesRegeneration of The LiverMonica Wyona LorensiaNo ratings yet
- Cell CycleDocument20 pagesCell CycleShelrenNo ratings yet
- Drosophila Eggshell Production - Identification of New Genes and Coordination by PXTDocument12 pagesDrosophila Eggshell Production - Identification of New Genes and Coordination by PXTFernandoNo ratings yet
- BIOLOGY OF REPRODUCTION 50, 765-773 (1994) : 'SupportedDocument9 pagesBIOLOGY OF REPRODUCTION 50, 765-773 (1994) : 'SupportedLateecka R KulkarniNo ratings yet
- CAP 2 Regulation of Follicle Formation andDocument27 pagesCAP 2 Regulation of Follicle Formation andgilson_pessoaNo ratings yet
- UntitledDocument234 pagesUntitledMarco Antonio AlvarezNo ratings yet
- Fisiologi Siklus Haid (DR - Kanadi SumaprajaDocument70 pagesFisiologi Siklus Haid (DR - Kanadi SumaprajaIntan DhyaksariNo ratings yet
- Oocyte Maturation: A Story of Arrest and Release: Frontiers in Bioscience (Scholar Edition) January 2013Document55 pagesOocyte Maturation: A Story of Arrest and Release: Frontiers in Bioscience (Scholar Edition) January 2013ArinilHaqueNo ratings yet
- It Takes Two To TangoDocument3 pagesIt Takes Two To Tangoaftab khanNo ratings yet
- Fisiologi HaidDocument66 pagesFisiologi HaidRizna AriyaniNo ratings yet
- Handouts Gen. Bio Oct 3 6 2022Document14 pagesHandouts Gen. Bio Oct 3 6 2022Mark Kenneth CastilloNo ratings yet
- Growth Factors and Growth Factor Receptors in Cancer: T. RajkumarDocument7 pagesGrowth Factors and Growth Factor Receptors in Cancer: T. RajkumarMohammed AladhraeiNo ratings yet
- Goats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationDocument11 pagesGoats Reproduction Estrual Cycle Estrous Synchronization Artificial InseminationBayu Anke MardianNo ratings yet
- Envejecimiento ReproductivoDocument10 pagesEnvejecimiento ReproductivoLAURA DANIELA VERA BELTRANNo ratings yet
- 6 Buratini 2012Document6 pages6 Buratini 2012Snery ZeusNo ratings yet
- Module 2. Topic 1 B2 Control of The Cell CycleDocument10 pagesModule 2. Topic 1 B2 Control of The Cell CycleRey Donn VILLABLANCANo ratings yet
- Wilson 2015Document8 pagesWilson 2015ngvy520201No ratings yet
- Book 2011Document102 pagesBook 2011fdsjklsdfxsdfsdfNo ratings yet
- Hormonal Regulation of Implantation: Pinar H. Kodaman, MD, PHD, Hugh S. Taylor, MDDocument22 pagesHormonal Regulation of Implantation: Pinar H. Kodaman, MD, PHD, Hugh S. Taylor, MDnurul hidayahNo ratings yet
- PCNA Marks The Initiation of Follicular Growth in The RatDocument8 pagesPCNA Marks The Initiation of Follicular Growth in The RatLinh Tran KhanhNo ratings yet
- AdsadasdadsadDocument1 pageAdsadasdadsadmihrullah.azimi1No ratings yet
- Role of Incretins in Pancreas Growth and Development: EditorialDocument3 pagesRole of Incretins in Pancreas Growth and Development: EditorialHarsha Ramakrishna KashyapNo ratings yet
- T10. Signaling in Control of Cell Growth and MetabolismDocument16 pagesT10. Signaling in Control of Cell Growth and MetabolismJavierNo ratings yet
- CellDocument16 pagesCellHilma HamidahNo ratings yet
- DNA Replication Chromosomes: Cell CycleDocument6 pagesDNA Replication Chromosomes: Cell CycleAnnisa FitriaNo ratings yet
- Developmental Human Leydig CellDocument9 pagesDevelopmental Human Leydig CellDelavemia RNo ratings yet
- Cell DeathDocument15 pagesCell DeathalbertoNo ratings yet
- Cell Cycle ApoptosisDocument17 pagesCell Cycle ApoptosisSuleiman DaudaNo ratings yet
- 2010 Vital Et AlDocument12 pages2010 Vital Et Alsbd2hugNo ratings yet
- Boden Steiner 1996Document14 pagesBoden Steiner 1996Nicolas Cabrera GimenaNo ratings yet
- Prolactin Modulates Steroidogenesis by Rat Granulosa Cells: I. Effects On Progesterone1Document8 pagesProlactin Modulates Steroidogenesis by Rat Granulosa Cells: I. Effects On Progesterone1Lateecka R KulkarniNo ratings yet
- Inc Prevalnce Retrospective Evidence 1994 PetersonDocument25 pagesInc Prevalnce Retrospective Evidence 1994 PetersonKurt SciclunaNo ratings yet
- Carcinogenesis - Arno HelmbergDocument4 pagesCarcinogenesis - Arno HelmbergHafsah ShoaibNo ratings yet
- Review Article: Molecular Mechanisms Responsible For Increased Vulnerability of The Ageing Oocyte To Oxidative DamageDocument22 pagesReview Article: Molecular Mechanisms Responsible For Increased Vulnerability of The Ageing Oocyte To Oxidative DamageokerNo ratings yet
- The Prevention of Aging in MammalsDocument5 pagesThe Prevention of Aging in MammalsJeremy KnippleNo ratings yet
- Glucocorticoid Metabolism in Equine Follicles and OocytesDocument12 pagesGlucocorticoid Metabolism in Equine Follicles and OocytesVerónica BecerraNo ratings yet
- Biología Del Cuerpo LuteoDocument12 pagesBiología Del Cuerpo LuteoVioleta v cNo ratings yet
- Molecular Mechanism of ImplantationDocument19 pagesMolecular Mechanism of ImplantationAbunu TilayeNo ratings yet
- Progress in Research On Key Factors Regulating Lactation. Guo, H., Li, J., Wang, Y., Cao, X., LV, X., Yang, Z., & Chen, Z. (2023) .Document11 pagesProgress in Research On Key Factors Regulating Lactation. Guo, H., Li, J., Wang, Y., Cao, X., LV, X., Yang, Z., & Chen, Z. (2023) .Nxyz GxyzNo ratings yet
- Superovulation and in Vitro Oocyte Maturation in Three SpeciesDocument10 pagesSuperovulation and in Vitro Oocyte Maturation in Three SpeciesAgustin Carmona CastroNo ratings yet
- Endocrine ReviewDocument9 pagesEndocrine ReviewPatty OlszynskiNo ratings yet
- Glycolysis & Cancer Write UpDocument9 pagesGlycolysis & Cancer Write UpMarvin JeaNo ratings yet
- Nutritional in Uences On Implantation and Placental DevelopmentDocument9 pagesNutritional in Uences On Implantation and Placental Developmentmustaring_mangkona8013No ratings yet
- Chapter+2Cell+Proliferation Onco CeliaDocument34 pagesChapter+2Cell+Proliferation Onco CeliaPaula C GuimaraesNo ratings yet
- Cow EstrusDocument4 pagesCow EstrusMageja TatendaNo ratings yet
- ID Pola Distribusi Mitokondria Sel Sel TrofDocument10 pagesID Pola Distribusi Mitokondria Sel Sel TrofRahmad ArfandiNo ratings yet
- A Cellular System That Degrades Misfolded Proteins and Protects Against Neurodeneration (Standart)Document16 pagesA Cellular System That Degrades Misfolded Proteins and Protects Against Neurodeneration (Standart)Andonis AngelovNo ratings yet
- Expression of p97/VCP (Valosin Containing Protein) and Jab1/CSN5 in Rat Testis and Epididymis During The Postnatal DevelopmentDocument26 pagesExpression of p97/VCP (Valosin Containing Protein) and Jab1/CSN5 in Rat Testis and Epididymis During The Postnatal DevelopmentdrmacrohardNo ratings yet
- Cell CycleDocument7 pagesCell CycleDr-Dalya ShakirNo ratings yet
- Evaluation of The Effects of CCN4 On Pancreatic Beta Cell ProliferationDocument18 pagesEvaluation of The Effects of CCN4 On Pancreatic Beta Cell ProliferationAriadna Holali MartínezNo ratings yet
- Stimulus-Triggered Fate Conversion of Somatic Cells Into PluripotencyDocument19 pagesStimulus-Triggered Fate Conversion of Somatic Cells Into PluripotencynembutalNo ratings yet
- The Ovarian ReserveDocument19 pagesThe Ovarian Reservepramod singhNo ratings yet
- Female HormoneDocument43 pagesFemale Hormoneامجد حسين جواد كاظمNo ratings yet
- Interaksi Endokrin Dalam Pengaturan Pertumbuhan JaninDocument40 pagesInteraksi Endokrin Dalam Pengaturan Pertumbuhan JaninFeni RiandariNo ratings yet
- Grimm 2020Document16 pagesGrimm 2020MD LarasatiNo ratings yet
- Molecular Regulation of Pancreatic Beta-Cell Mass DevelopmentDocument14 pagesMolecular Regulation of Pancreatic Beta-Cell Mass DevelopmentAndrei MitescuNo ratings yet
- Regenerative Medicine in Diabetes: General IntroductionDocument10 pagesRegenerative Medicine in Diabetes: General IntroductionAlex PlicNo ratings yet
- Principles of Gas Chromatography 2Document12 pagesPrinciples of Gas Chromatography 2Enanahmed EnanNo ratings yet
- Florinsky 2010dDocument403 pagesFlorinsky 2010dVostjeNo ratings yet
- NeoplasiaDocument15 pagesNeoplasiaAmir Shafiq100% (1)
- LP9th - Cardiovascular SystemDocument5 pagesLP9th - Cardiovascular SystemOliver VillanuevaNo ratings yet
- Integrated Science NotesDocument17 pagesIntegrated Science NotesannmarieNo ratings yet
- Bio RevisionsheetDocument4 pagesBio RevisionsheetKhushbu DaveyNo ratings yet
- Lecture Histology USDocument7 pagesLecture Histology USالهاشميNo ratings yet
- IntelliegenceDocument31 pagesIntelliegenceasha suratkalNo ratings yet
- Structure and Functions of The EyeDocument16 pagesStructure and Functions of The EyeabdirizakNo ratings yet
- Blakemore and CooperDocument2 pagesBlakemore and Coopermeenaxi.aravinthanNo ratings yet
- Topic 1 1-24-22 - UploadDocument152 pagesTopic 1 1-24-22 - UploadGloria VasquezNo ratings yet
- The Ultimate Compost Tea Guide - KIS OrganicsDocument16 pagesThe Ultimate Compost Tea Guide - KIS OrganicsakshayNo ratings yet
- Reduction of Organic Load and Biodegradation of PaDocument19 pagesReduction of Organic Load and Biodegradation of PaArie Ikhwan SaputraNo ratings yet
- Stages of MitosisDocument2 pagesStages of MitosisWONGNo ratings yet
- Contrast Medium Experiment With A Blood Vessel ModelDocument3 pagesContrast Medium Experiment With A Blood Vessel ModelJose GalvanNo ratings yet
- Smooth MuscleDocument27 pagesSmooth MuscleMichael GodwinNo ratings yet
- Ultrasound DiathermiDocument1 pageUltrasound DiathermiPatrick MuljonoNo ratings yet
- Janka Wood Hardness RatingDocument3 pagesJanka Wood Hardness Ratingsaiga3089417No ratings yet
- Name: Nornisah H. Pangandaman Section: BSN 2A Fdar Date & Time Focus Data Action ResponseDocument2 pagesName: Nornisah H. Pangandaman Section: BSN 2A Fdar Date & Time Focus Data Action ResponseNornisah H. PangandamanNo ratings yet
- Auditory SystemDocument8 pagesAuditory SystemRox SanNo ratings yet
- DR Divyesh ISOT 2023Document64 pagesDR Divyesh ISOT 2023divyesh engineerNo ratings yet
- MDR Tecnical Sheet With Digital SignatureDocument103 pagesMDR Tecnical Sheet With Digital SignatureMamdouhNo ratings yet
- Educ 121 Theorist Presentation Urie BronfenbrennerDocument13 pagesEduc 121 Theorist Presentation Urie Bronfenbrennerapi-285371570No ratings yet
- Biomechanics of MusclesDocument58 pagesBiomechanics of MusclesAsad Chaudhary100% (2)
- Jurnal Paliatif 1Document10 pagesJurnal Paliatif 1Anisa SilviaNo ratings yet
- Isolation of Plasmid DNA From Agrobacterium Using The QIAprep® Spin Miniprep Kit Spin ProcedureDocument3 pagesIsolation of Plasmid DNA From Agrobacterium Using The QIAprep® Spin Miniprep Kit Spin Procedure于连升No ratings yet
- Tropical Deciduous Forest: GeographyDocument12 pagesTropical Deciduous Forest: GeographyPUJA THAKURNo ratings yet
- Connecting Chemistry To Our WorldDocument8 pagesConnecting Chemistry To Our WorldjyclynnnNo ratings yet
- Phosphate DeterminationDocument5 pagesPhosphate DeterminationCaryl FrancheteNo ratings yet
- DSFSFDocument3 pagesDSFSFruchelNo ratings yet