Download as pdf or txt
You might also like
- ABAS II Parent (0 5) Sample Test ReportDocument7 pagesABAS II Parent (0 5) Sample Test ReportHKNo ratings yet
- Kotler Pom16e Inppt 05Document48 pagesKotler Pom16e Inppt 05gosaye desalegn100% (2)
- Forms of CommunicationDocument12 pagesForms of Communicationdominic laroza50% (2)
- Udl Lesson Plan Form Rev1Document3 pagesUdl Lesson Plan Form Rev1api-239644736No ratings yet
- 2) Cui (2017)Document30 pages2) Cui (2017)Êndel AlvesNo ratings yet
- 2009 Kang - Insulin Secreting Cells From Human Eyelid Derived Stem Cells Alleviate Type I Diabetes in Immunocompetent MiceDocument10 pages2009 Kang - Insulin Secreting Cells From Human Eyelid Derived Stem Cells Alleviate Type I Diabetes in Immunocompetent MiceNyna KawlesNo ratings yet
- Anucleate Platelets Generate Progeny 1Document34 pagesAnucleate Platelets Generate Progeny 1Alejandro BarretoNo ratings yet
- Effect of Mesenchymal Stem Cells Transplantation On Glycaemic Profile & Their Localization in Streptozotocin Induced Diabetic Wistar RatsDocument9 pagesEffect of Mesenchymal Stem Cells Transplantation On Glycaemic Profile & Their Localization in Streptozotocin Induced Diabetic Wistar RatsShanaz NovriandinaNo ratings yet
- Yi - Et - Al-Glia 2017Document19 pagesYi - Et - Al-Glia 2017Daniela HenríquezNo ratings yet
- Ehler201dd7 Article TranslationalEvidenceForTwoDisDocument10 pagesEhler201dd7 Article TranslationalEvidenceForTwoDisMark TomNo ratings yet
- 1 s2.0 S0361923016304105 MainDocument5 pages1 s2.0 S0361923016304105 MainMKGUNo ratings yet
- MSC Reduces Il-6 Gene Expression in AdDocument16 pagesMSC Reduces Il-6 Gene Expression in AdFarhan Royan PermanahadiNo ratings yet
- Metabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDocument12 pagesMetabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDavid Galiano LatorreNo ratings yet
- Lactate A Neuro SourceDocument9 pagesLactate A Neuro SourceDaniel Lee Eisenberg JacobsNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument7 pagesBiochemical and Biophysical Research CommunicationsGiulia AndreeaNo ratings yet
- Blood Cells, Molecules and Diseases: SciencedirectDocument14 pagesBlood Cells, Molecules and Diseases: SciencedirectdrhiteshchopraNo ratings yet
- J Cell Sci-1993-Perou-99-107Document9 pagesJ Cell Sci-1993-Perou-99-107glodovichiNo ratings yet
- BBB & EpilepsyDocument14 pagesBBB & EpilepsyBujoreanu IulianNo ratings yet
- Awab 134Document12 pagesAwab 134yi1zhangNo ratings yet
- Stemcells 2006-0409Document9 pagesStemcells 2006-0409dupuytrenNo ratings yet
- Lee 2008Document14 pagesLee 2008Laura Duarte RojasNo ratings yet
- Bone Marrow Mastocytosis in Dogs With Myelosuppressive Monocytic Ehrlichiosis (Ehrlichia Canis) : A Retrospective StudyDocument4 pagesBone Marrow Mastocytosis in Dogs With Myelosuppressive Monocytic Ehrlichiosis (Ehrlichia Canis) : A Retrospective StudyDiana GranadaNo ratings yet
- Endotelial Pericites - Cui2006Document10 pagesEndotelial Pericites - Cui2006Lilo FrancoisNo ratings yet
- Endothelial Cell NADPH Oxidase Mediates The Cerebral Microvascular Dysfunction in Sickle Cell Transgenic MiceDocument19 pagesEndothelial Cell NADPH Oxidase Mediates The Cerebral Microvascular Dysfunction in Sickle Cell Transgenic MiceNovia MentariNo ratings yet
- Cerebrolysin Increases Recovery and Decreases Inflammation in An Autoimmune Model of Autoimmune EncephalitisDocument15 pagesCerebrolysin Increases Recovery and Decreases Inflammation in An Autoimmune Model of Autoimmune EncephalitisAlin MihaiNo ratings yet
- Axon-Glia Synapses Are Highly Vulnerable To White Matter Injury in The Developing BrainDocument17 pagesAxon-Glia Synapses Are Highly Vulnerable To White Matter Injury in The Developing BrainKing Man LoNo ratings yet
- Citosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Document1 pageCitosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Alondra MaldonadoNo ratings yet
- DM Tipe 1Document6 pagesDM Tipe 1Mutiara Kristiani PutriNo ratings yet
- Rapid and Efficient Generation of OligodendrocytesDocument22 pagesRapid and Efficient Generation of Oligodendrocytessabahmozafari21No ratings yet
- 2017 Article 199Document13 pages2017 Article 199Rabiatul 'raney' AdawiyahNo ratings yet
- Takenouchi 20151Document13 pagesTakenouchi 20151Angelique R. AltamiranoNo ratings yet
- Biochem Journal CritiqueDocument7 pagesBiochem Journal CritiqueDiane TomogbongNo ratings yet
- Recuperarea Defectului de Dimensiune Critică Al Osului Parietal La Iepure Prin Grefe CompoziteDocument7 pagesRecuperarea Defectului de Dimensiune Critică Al Osului Parietal La Iepure Prin Grefe CompoziteRoxana VoicuNo ratings yet
- 06-Cryopreservation of Stem CellsDocument29 pages06-Cryopreservation of Stem Cellsjashuj.2004No ratings yet
- JCI2003Document8 pagesJCI2003kiwelem698No ratings yet
- Acido ValproicoDocument10 pagesAcido ValproicopreedtooneNo ratings yet
- Lund 2006Document17 pagesLund 2006marialuizacdasilvaNo ratings yet
- Xin 2013Document5 pagesXin 2013Maximiliano PfannlNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument7 pagesBiochemical and Biophysical Research CommunicationsAnthony CaldwellNo ratings yet
- Review Journal Experimental Model of Myocardial Infarction Induced by Isoproterenol in RatsDocument5 pagesReview Journal Experimental Model of Myocardial Infarction Induced by Isoproterenol in RatsM.Rizky irawanNo ratings yet
- Vitrification Preserves SSCS, Wyns 2013 PDFDocument12 pagesVitrification Preserves SSCS, Wyns 2013 PDFVarawenNo ratings yet
- Production of Functional Glucagon-Secreting From Human Embryonic Stem CellsDocument9 pagesProduction of Functional Glucagon-Secreting From Human Embryonic Stem CellsNyna KawlesNo ratings yet
- Effects of Diabetes On Apoptosis and Mitosis in Rat HippocampusDocument9 pagesEffects of Diabetes On Apoptosis and Mitosis in Rat HippocampusFrancelia Quiñonez RuvalcabaNo ratings yet
- Atiakshin 2017Document12 pagesAtiakshin 2017iuventasNo ratings yet
- Differentiation of Human Multipotent Dermal Fibroblasts Into Islet-Like Cell ClustersDocument8 pagesDifferentiation of Human Multipotent Dermal Fibroblasts Into Islet-Like Cell Clustersanil0811No ratings yet
- Research Open Access: Charlotte Schmitt, Nathalie Strazielle and Jean-François Ghersi-EgeaDocument15 pagesResearch Open Access: Charlotte Schmitt, Nathalie Strazielle and Jean-François Ghersi-Egeamihail.boldeanuNo ratings yet
- Diabetes 2005 Carrillo 69 77Document9 pagesDiabetes 2005 Carrillo 69 77Grigorina MitrofanNo ratings yet
- Short Term Induction of Lps Damages The Hippocampus Gyrus Through Apoptosis On Glial Cells MiceDocument9 pagesShort Term Induction of Lps Damages The Hippocampus Gyrus Through Apoptosis On Glial Cells MiceGlobal Research and Development ServicesNo ratings yet
- 0007 0012 PDFDocument6 pages0007 0012 PDFHalim SudonoNo ratings yet
- Antibody Appears To Attack Cancer Cells, Leaving Other Cells UnscathedDocument13 pagesAntibody Appears To Attack Cancer Cells, Leaving Other Cells UnscathedAnonymous EdgflnjcNo ratings yet
- Immortalized Liver Endothel CCmodelDocument20 pagesImmortalized Liver Endothel CCmodelAndreiNo ratings yet
- Association of Mast CellsDocument8 pagesAssociation of Mast CellsJéssica MacêdoNo ratings yet
- Phil. Trans. R. Soc. B 2011 Unternaehrer 2274 85Document13 pagesPhil. Trans. R. Soc. B 2011 Unternaehrer 2274 85Jo-Anne M. VentiganNo ratings yet
- Increased Islet Apoptosis in Pdx1 MiceDocument14 pagesIncreased Islet Apoptosis in Pdx1 Miceroh009No ratings yet
- Leukocyte Survival in Cerebrospinal FluidDocument2 pagesLeukocyte Survival in Cerebrospinal FluidntnquynhproNo ratings yet
- Transplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Document128 pagesTransplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Enrique Moreno GonzálezNo ratings yet
- Estudio Histopatológico e Inmunohistoquímico de Páncreas en Perros Diabéticos Inducidos Con AloxanoDocument10 pagesEstudio Histopatológico e Inmunohistoquímico de Páncreas en Perros Diabéticos Inducidos Con AloxanoMiguel Chimal MuñozNo ratings yet
- Stemcell For Uni PakuanDocument71 pagesStemcell For Uni PakuanZaldyRusliNo ratings yet
- Induced Monocytes-Derived Hscs (Cd34+) With Lps Accelerated Homing Rat Bone Marrow-Mesenchymal Stem Cell (BM-MSCS, Cd105) in Injured PancreasDocument12 pagesInduced Monocytes-Derived Hscs (Cd34+) With Lps Accelerated Homing Rat Bone Marrow-Mesenchymal Stem Cell (BM-MSCS, Cd105) in Injured PancreasAzmi FarhadiNo ratings yet
- A Novel, Highly Sensitive and Specific Biomarker For Niemann-Pick Type C1 DiseaseDocument8 pagesA Novel, Highly Sensitive and Specific Biomarker For Niemann-Pick Type C1 DiseaseTheodora TeddyNo ratings yet
- Thermogenic Activity of UCP1 in Human White Fat-Derived Beige Adipocytes 2015 ResearchDocument10 pagesThermogenic Activity of UCP1 in Human White Fat-Derived Beige Adipocytes 2015 Research畏No ratings yet
- Metabolites 09 00013Document15 pagesMetabolites 09 00013Masoome ANo ratings yet
- A Comparison of Immunohistochemical and Silver Staining Methods For The Detection of Diffuse Plaques ... (2006)Document13 pagesA Comparison of Immunohistochemical and Silver Staining Methods For The Detection of Diffuse Plaques ... (2006)Apróné Török IbolyaNo ratings yet
- The Nucleus Basalis of Meynert in Multi-Infarct (Vascular) DementiaDocument6 pagesThe Nucleus Basalis of Meynert in Multi-Infarct (Vascular) Dementiadimitri pizarnikNo ratings yet
- Effects of Zizyphus Jujube Extract On Memory and Learning Impairment Induced by Bilateral Electric Lesions of The Nucleus Basalis of Meynert in RatDocument8 pagesEffects of Zizyphus Jujube Extract On Memory and Learning Impairment Induced by Bilateral Electric Lesions of The Nucleus Basalis of Meynert in Ratdimitri pizarnikNo ratings yet
- Alzheimer'S Disease and Their Relationship To Ageing and To TheDocument16 pagesAlzheimer'S Disease and Their Relationship To Ageing and To Thedimitri pizarnikNo ratings yet
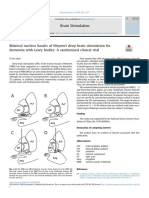
- Bilateral Nucleus Basalis of Meynert Deep Brain Stimulation For Dementia With Lewy Bodies - A Randomised Clinical TrialDocument2 pagesBilateral Nucleus Basalis of Meynert Deep Brain Stimulation For Dementia With Lewy Bodies - A Randomised Clinical Trialdimitri pizarnikNo ratings yet
- Chapter 04 The Self, Identity, Emotion, and PersonalityDocument22 pagesChapter 04 The Self, Identity, Emotion, and PersonalityTracy AndersonNo ratings yet
- Severe DisabilitiesDocument10 pagesSevere DisabilitiesRazelle Angiela AvorqueNo ratings yet
- HTH SCI 4AA3 at McMaster U, RolesDocument5 pagesHTH SCI 4AA3 at McMaster U, Rolesstargaze_night_42No ratings yet
- The Use of Photos in Making Reflective Essays (UPDATED)Document17 pagesThe Use of Photos in Making Reflective Essays (UPDATED)Jazel Mabilanga RobleNo ratings yet
- Embracing The Whold Gifted Self - July19 Sample Pages 1 20Document24 pagesEmbracing The Whold Gifted Self - July19 Sample Pages 1 20Erick GüitrónNo ratings yet
- Books Are Better Than TVDocument10 pagesBooks Are Better Than TVMohan updykadirNo ratings yet
- Dream RecallDocument2 pagesDream RecallGabriel MarinNo ratings yet
- Self Inquiry GuideDocument15 pagesSelf Inquiry GuideRolf KrahnertNo ratings yet
- 4 - Artificial Intelligence in Neurodegenerative Diseases A Review of Available Tools With A Focus On Machine Learning TechniquesDocument22 pages4 - Artificial Intelligence in Neurodegenerative Diseases A Review of Available Tools With A Focus On Machine Learning TechniquesGUILLERMO PRIETO AVALOSNo ratings yet
- Trauma and Dissociation - Implications For Borderline Personality DisorderDocument10 pagesTrauma and Dissociation - Implications For Borderline Personality DisorderCarla MesquitaNo ratings yet
- Biological Basis of BehaviourDocument6 pagesBiological Basis of BehaviourRichard ManuelNo ratings yet
- English Lang AnxDocument15 pagesEnglish Lang AnxAngela ArdidonNo ratings yet
- Emotional Intelligence-Ayo Maria OdusanyaDocument28 pagesEmotional Intelligence-Ayo Maria OdusanyadeeNo ratings yet
- Time BoxDocument4 pagesTime BoxLaster Jauod100% (2)
- Career DevelopmentDocument26 pagesCareer DevelopmentAngeline ParadoNo ratings yet
- Reading Comprehension and Mathematical Performance in Solving Word Problems Among Grade 4 LearnersDocument9 pagesReading Comprehension and Mathematical Performance in Solving Word Problems Among Grade 4 LearnersPsychology and Education: A Multidisciplinary JournalNo ratings yet
- Module 2 CompilationDocument25 pagesModule 2 CompilationKathleen ContapayNo ratings yet
- Pidato Bahasa Inggris Dan Indonesia Tentang CitaDocument2 pagesPidato Bahasa Inggris Dan Indonesia Tentang CitaMIFTAHUL ULUM100% (1)
- 1995 Pallarés Nadal Ketamine Lever PessDocument4 pages1995 Pallarés Nadal Ketamine Lever PessRocío Donaire CortésNo ratings yet
- PostureDocument46 pagesPostureFarrukh Shahzad100% (1)
- Design Thinking-3Document45 pagesDesign Thinking-3Joel JeremyNo ratings yet
- (C) Trait (D) None of The AboveDocument12 pages(C) Trait (D) None of The AboveTriyogaAgungNo ratings yet
- 12btypes of Postural ReflexesDocument1 page12btypes of Postural ReflexesShaleen SinghNo ratings yet
- 6 Cognitive Behavioral Therapy Techniques For Bipolar DisorderDocument7 pages6 Cognitive Behavioral Therapy Techniques For Bipolar Disorderprabha50% (2)
- Utbk 3 Ppu PembahasanDocument16 pagesUtbk 3 Ppu PembahasanJuita Ilda yaniNo ratings yet
- Introduction To Mental Health and Hygiene and Adjustment MechanismDocument18 pagesIntroduction To Mental Health and Hygiene and Adjustment MechanismLakshmiRengarajan94% (17)