
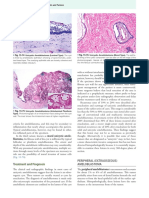
Resveratrol Alleviates Temporomandibular Joint Inflammatory Pain by Recovering Disturbed Gut Microbiota
Resveratrol Alleviates Temporomandibular Joint Inflammatory Pain by Recovering Disturbed Gut Microbiota
You might also like
- 057608komatsu Hydraulic Oil HO 46-HMDocument5 pages057608komatsu Hydraulic Oil HO 46-HMrolex21No ratings yet
- 601 2023 30Document8 pages601 2023 30Shahram AwanNo ratings yet
- s10787 017 0311 XDocument4 pagess10787 017 0311 X2nmjtzn9rgNo ratings yet
- 13 - BPS Update TOG 2022Document8 pages13 - BPS Update TOG 2022Anna JuniedNo ratings yet
- Gut Microbiota in Neurodegenerative DisordersDocument31 pagesGut Microbiota in Neurodegenerative DisordersMércia FiuzaNo ratings yet
- Isola Etal 2020 Clo IDocument11 pagesIsola Etal 2020 Clo IKAREN ESTHER CAPACOILA GRANADOSNo ratings yet
- Histamine H2 Receptor-Mediated Suppression of Intestinal Inflammation by Probiotic Lactobacillus ReuteriDocument12 pagesHistamine H2 Receptor-Mediated Suppression of Intestinal Inflammation by Probiotic Lactobacillus ReuteriNejc KovačNo ratings yet
- The Role of Prophylactic Ibuprofen and N-Acetylcysteine On The Level of Cytokines in Periapical Exudates and The Post-Treatment PainDocument6 pagesThe Role of Prophylactic Ibuprofen and N-Acetylcysteine On The Level of Cytokines in Periapical Exudates and The Post-Treatment PainNadya PurwantyNo ratings yet
- Article 180803Document2 pagesArticle 180803mustafaali9711No ratings yet
- Review: Mind-Altering With The Gut: Modulation of The Gut-Brain Axis With ProbioticsDocument11 pagesReview: Mind-Altering With The Gut: Modulation of The Gut-Brain Axis With ProbioticsVitor PereiraNo ratings yet
- Fcimb 12 812596Document20 pagesFcimb 12 812596andreafernandez2eNo ratings yet
- Spielman Et Al., 2018-Unhealthy Gut, Unhealthy Brain - The Role of The Intestinal Microbiota in Neurodegenerative DiseasesDocument15 pagesSpielman Et Al., 2018-Unhealthy Gut, Unhealthy Brain - The Role of The Intestinal Microbiota in Neurodegenerative DiseasesEdwar MagallanNo ratings yet
- Journal of Herbal MedicineDocument8 pagesJournal of Herbal MedicineCarlos Andrés Gallardo GarridoNo ratings yet
- Akinyemi Emmanuel ifeoluwa IBD (2)-1-1Document45 pagesAkinyemi Emmanuel ifeoluwa IBD (2)-1-1eakinyemi360No ratings yet
- Mechanisms of Peptic Ulcer Recurrence: Role of in Ammation: ReviewDocument12 pagesMechanisms of Peptic Ulcer Recurrence: Role of in Ammation: ReviewGuilherme Vieira Botelho de Almeida (Teacher Red Balloon Cuiabá)No ratings yet
- The Gut Microbiota in Anxiety and Depression - A Systematic Review (Simpson Et Al, 2020)Document18 pagesThe Gut Microbiota in Anxiety and Depression - A Systematic Review (Simpson Et Al, 2020)GokushimakNo ratings yet
- Akinyemi Emmanuel ifeoluwa IBD (2)-1Document46 pagesAkinyemi Emmanuel ifeoluwa IBD (2)-1eakinyemi360No ratings yet
- 13. MicroRNAs 106b and 222 Improve Hyperglycemia in a Mouse Model of Insulin - Deficient Diabetes via Pancreatic Β-Cell ProliferationDocument10 pages13. MicroRNAs 106b and 222 Improve Hyperglycemia in a Mouse Model of Insulin - Deficient Diabetes via Pancreatic Β-Cell ProliferationvirginiaviolaNo ratings yet
- The_microbiota-gut-brain_axis_regulates_motivationDocument3 pagesThe_microbiota-gut-brain_axis_regulates_motivationchaiyadol tantasithNo ratings yet
- Botulinum Toxin in CancerDocument11 pagesBotulinum Toxin in CancerAlicia Ramirez HernandezNo ratings yet
- Host Modulation Therapy An Updated ReviewDocument4 pagesHost Modulation Therapy An Updated Reviewpaper kitaNo ratings yet
- The Anti Inflammatory Effect of Triphala in Arthritic Induced RatsDocument11 pagesThe Anti Inflammatory Effect of Triphala in Arthritic Induced RatsShyam S SundarNo ratings yet
- Bethany Pinnock - Periodontics V Research Paper (Final)Document5 pagesBethany Pinnock - Periodontics V Research Paper (Final)Bethany PinnockNo ratings yet
- 80 BiotaDocument7 pages80 BiotaAlfonso De la FuenteNo ratings yet
- Raja Et Al 2013Document7 pagesRaja Et Al 2013Sahera Nurhidayah NasutionNo ratings yet
- Potential Influences of Gut Microbiota On The Formation of Intracranial AneurysmDocument14 pagesPotential Influences of Gut Microbiota On The Formation of Intracranial AneurysmMércia FiuzaNo ratings yet
- The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain CommunicationDocument14 pagesThe Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain CommunicationMércia FiuzaNo ratings yet
- Diet MS Valter LongoDocument12 pagesDiet MS Valter LongoAnonymous 0qYv7WpgNo ratings yet
- 1 s2.0 S0163725822002212 MainDocument29 pages1 s2.0 S0163725822002212 MainNélida Lucia Rodriguez AscañoNo ratings yet
- Jurnal SR 3Document12 pagesJurnal SR 3Zaina Khairunisa AsyaNo ratings yet
- Manzhulo 2015Document8 pagesManzhulo 2015Yolla JufandaNo ratings yet
- Animal Models of Rheumatoid Pain: Experimental Systems and InsightsDocument12 pagesAnimal Models of Rheumatoid Pain: Experimental Systems and InsightsLourdes MendietaNo ratings yet
- 10.1515revneuro 2019 0005Document12 pages10.1515revneuro 2019 0005Suhartini RahmanNo ratings yet
- Probiotics Article 2019 PDFDocument10 pagesProbiotics Article 2019 PDFLuiis LimaNo ratings yet
- Share 10.1.1.825.5698Document36 pagesShare 10.1.1.825.5698Aidil CakepNo ratings yet
- Presentation Gut Brain August 2018Document17 pagesPresentation Gut Brain August 2018Sribharathi Indrajith100% (1)
- Tratament Conservator DupDocument3 pagesTratament Conservator DupDora PopaNo ratings yet
- Role of The Gut Microbiota in Ischemic Stroke: Katarzyna Winek, Ulrich Dirnagl, Andreas MeiselDocument7 pagesRole of The Gut Microbiota in Ischemic Stroke: Katarzyna Winek, Ulrich Dirnagl, Andreas MeiselArdianNo ratings yet
- Host Modulation in PeriodonticsDocument12 pagesHost Modulation in PeriodonticsFirma Nurdinia DewiNo ratings yet
- Keaa 192Document11 pagesKeaa 192germanNo ratings yet
- Inflammatory Bowel Disease DayaDocument74 pagesInflammatory Bowel Disease DayarajhiniNo ratings yet
- Role - and - Mechanism - of - Gut MicrobiomaDocument12 pagesRole - and - Mechanism - of - Gut MicrobiomaAyushi MauryaNo ratings yet
- Gut Dysbiosis, Leaky Gut, and Intestinal Epithelial Proliferation in Neurological Disorders Towards The Development of A New Therapeutic Using Amino Acids, Prebiotics, Probiotics, and PostbioticsDocument23 pagesGut Dysbiosis, Leaky Gut, and Intestinal Epithelial Proliferation in Neurological Disorders Towards The Development of A New Therapeutic Using Amino Acids, Prebiotics, Probiotics, and PostbioticsLevente BalázsNo ratings yet
- Nutrients 13 03192 v3Document17 pagesNutrients 13 03192 v3Firman Kusuma AtmajaNo ratings yet
- The Gut Microbiota in Inflammatory Bowel DiseaseDocument14 pagesThe Gut Microbiota in Inflammatory Bowel DiseaseMércia FiuzaNo ratings yet
- Jurnal 3Document20 pagesJurnal 3mardha tillahNo ratings yet
- 2020 - Guo - Oligosaccharadide TMVDocument7 pages2020 - Guo - Oligosaccharadide TMVandreeboneNo ratings yet
- Gastroprotectants in Small Animal Veterinary Practice - A Review of The Evidence Part 1 - Cyto-Protective DrugsDocument16 pagesGastroprotectants in Small Animal Veterinary Practice - A Review of The Evidence Part 1 - Cyto-Protective DrugsPatriciaNo ratings yet
- Molecules 28 00106Document17 pagesMolecules 28 00106Jeoval NetoNo ratings yet
- Lect 1 Host ModulationDocument22 pagesLect 1 Host ModulationrupaliNo ratings yet
- Botox DTM Neuralgia V Par EtcDocument16 pagesBotox DTM Neuralgia V Par EtcjosfranNo ratings yet
- Bromelain A Potential Strategy For The Adjuvant Treatment of PeriodontitisDocument6 pagesBromelain A Potential Strategy For The Adjuvant Treatment of Periodontitismajiamal99No ratings yet
- Article 1 Irfan KhanDocument19 pagesArticle 1 Irfan KhanAshique RajputNo ratings yet
- Format Jurnal Pak Kir 141018Document7 pagesFormat Jurnal Pak Kir 141018daniel_alexander_susenoNo ratings yet
- Cranio®: The Journal of Craniomandibular & Sleep PracticeDocument11 pagesCranio®: The Journal of Craniomandibular & Sleep PracticeAnthony VillafañeNo ratings yet
- Nsaids in PeriodontologyDocument7 pagesNsaids in Periodontologysalman khawarNo ratings yet
- Diacerein New CliicalDocument9 pagesDiacerein New Cliicaledo adimastaNo ratings yet
- Probiotic Antigenotoxic Activity As A DNA Bioprotective Tool: A Minireview With Focus On Endocrine DisruptorsDocument11 pagesProbiotic Antigenotoxic Activity As A DNA Bioprotective Tool: A Minireview With Focus On Endocrine DisruptorsAjit Kumar BaidyaNo ratings yet
- Messengers From The MicrobiotaDocument3 pagesMessengers From The MicrobiotaDra. Ianna CarmoNo ratings yet
- Oral Probiotic Treatment of Lactobacillus Rhamnosus Lcr35® Prevents Visceral Hypersensitivity To A Colonic Inflammation and An Acute Psychological StressDocument13 pagesOral Probiotic Treatment of Lactobacillus Rhamnosus Lcr35® Prevents Visceral Hypersensitivity To A Colonic Inflammation and An Acute Psychological StressMaria TejedaNo ratings yet
- The new frontier of anti-aging medicine: Miracles nutrigenomics senolitic products for a healthy lifeFrom EverandThe new frontier of anti-aging medicine: Miracles nutrigenomics senolitic products for a healthy lifeNo ratings yet
- jcdr-9-ZE08Document4 pagesjcdr-9-ZE08germanNo ratings yet
- The Use of Pharmacologic Agents in The Management of Temporomandibular Joint DisorderDocument7 pagesThe Use of Pharmacologic Agents in The Management of Temporomandibular Joint DisordergermanNo ratings yet
- The Problems Preventing Progress in Adhesive Restorative DentistryDocument6 pagesThe Problems Preventing Progress in Adhesive Restorative DentistrygermanNo ratings yet
- Ribeiro 2020Document8 pagesRibeiro 2020germanNo ratings yet
- Keaa 192Document11 pagesKeaa 192germanNo ratings yet
- Meta 2.1 Ocampo Libro (15-21)Document7 pagesMeta 2.1 Ocampo Libro (15-21)germanNo ratings yet
- Meta 2.1 Ocampo LibroDocument21 pagesMeta 2.1 Ocampo LibrogermanNo ratings yet
- Meta 2.1 Ocampo Libro (08-14)Document7 pagesMeta 2.1 Ocampo Libro (08-14)germanNo ratings yet
- Meta 2.1 Ocampo Libro (01-07)Document7 pagesMeta 2.1 Ocampo Libro (01-07)germanNo ratings yet
- Meta 1.1 Ocampo Libro (11-21)Document11 pagesMeta 1.1 Ocampo Libro (11-21)germanNo ratings yet
- Meta 1.1 Ocampo Libro (11-21) (01-06)Document6 pagesMeta 1.1 Ocampo Libro (11-21) (01-06)germanNo ratings yet
- Meta 2.2 Ocampo LibroDocument7 pagesMeta 2.2 Ocampo LibrogermanNo ratings yet
- "After All, There Is Nothing As Interesting As People, and One Can Never Study Them Enough" Vincent Van GoghDocument38 pages"After All, There Is Nothing As Interesting As People, and One Can Never Study Them Enough" Vincent Van GoghAnonymous 1gH7ra9ANo ratings yet
- Hyperworks Technology Conference (HTC) 2011: Optimization of PMH StructuresDocument25 pagesHyperworks Technology Conference (HTC) 2011: Optimization of PMH StructuresgiakovidNo ratings yet
- Stresses and Challenges For New Graduates in HospitalsDocument6 pagesStresses and Challenges For New Graduates in HospitalskitsilNo ratings yet
- Omonigbehin Emmanuel AdedayoDocument12 pagesOmonigbehin Emmanuel AdedayoAriel GhieNo ratings yet
- 5X5 Intermediate - Bill StarrDocument7 pages5X5 Intermediate - Bill StarrAfrican MastiffNo ratings yet
- Which of The Following Is NOT True About Dust?Document3 pagesWhich of The Following Is NOT True About Dust?lucel palaca100% (1)
- Coriander (Corindrum Sativum L) and Its Bioactive ConstituentsDocument19 pagesCoriander (Corindrum Sativum L) and Its Bioactive ConstituentsFirda AuliyaNo ratings yet
- 10besar Kode PenyakitDocument5 pages10besar Kode PenyakitDEWINo ratings yet
- Closed PTW AuditDocument1 pageClosed PTW Auditf.BNo ratings yet
- Benefits of Hypnosis Author Mental Health America of Northern KentuckyDocument4 pagesBenefits of Hypnosis Author Mental Health America of Northern KentuckyJakarta SelamatNo ratings yet
- Health Quiz No. 2: NAME: - GRADE 6 - SCOREDocument2 pagesHealth Quiz No. 2: NAME: - GRADE 6 - SCORELove LeiNo ratings yet
- Hope - 1 Grade 11: Exercise For FitnessDocument6 pagesHope - 1 Grade 11: Exercise For FitnessTABBAL JAN PAOLONo ratings yet
- Margaret Paul - Taking The Lid Off Your FeelingsDocument4 pagesMargaret Paul - Taking The Lid Off Your FeelingspapetologeneNo ratings yet
- Medical Condition Report Free PDF TemplateDocument2 pagesMedical Condition Report Free PDF Templateapi-340236228No ratings yet
- WEEK 2: 12A, 12B, 12G Ôn Tập Khảo Sát Pronunciation ClosestDocument6 pagesWEEK 2: 12A, 12B, 12G Ôn Tập Khảo Sát Pronunciation ClosestHàNhậtNguyễnNo ratings yet
- Development of Push-Pull Osmotic Pump Tablets For A SlightlyDocument4 pagesDevelopment of Push-Pull Osmotic Pump Tablets For A SlightlyphamuyenthuNo ratings yet
- Life Calling Map Fillable FormDocument2 pagesLife Calling Map Fillable Formapi-300853489No ratings yet
- The Effects of Emotion On Short-Term Power Spectrum Analysis of Heart RateDocument5 pagesThe Effects of Emotion On Short-Term Power Spectrum Analysis of Heart RateManuel Guerrero GómezNo ratings yet
- Anna Garcia Death Timeline 1Document19 pagesAnna Garcia Death Timeline 1api-245176779No ratings yet
- Rose Doe v. The City of New York, Et. Al.Document57 pagesRose Doe v. The City of New York, Et. Al.Daily Caller News FoundationNo ratings yet
- Journal Medicine: The New EnglandDocument7 pagesJournal Medicine: The New EnglandTony RamirezNo ratings yet
- Knowledge ManagementDocument63 pagesKnowledge Managementrahul-singh-6592No ratings yet
- National Program For Rehabilitation of Polluted Site - A Case StudyDocument4 pagesNational Program For Rehabilitation of Polluted Site - A Case StudyijeteeditorNo ratings yet
- PREGNANCYDocument4 pagesPREGNANCYShaileeNo ratings yet
- Daftar Obat %pareto Dan Ven 15Document200 pagesDaftar Obat %pareto Dan Ven 15sofia nofiantiNo ratings yet
- UP Vaccine Center ListDocument106 pagesUP Vaccine Center ListRamendra Nath JhaNo ratings yet
- Language Disorders - 5Document27 pagesLanguage Disorders - 5Marta Sampedro GonzalezNo ratings yet
- Henares Vs LTFRBDocument2 pagesHenares Vs LTFRBCarolina Lim GambanNo ratings yet
- Maternity Nursing ReviewDocument37 pagesMaternity Nursing Reviewshenric16No ratings yet

































































































Download as pdf or txt
You might also like
- 057608komatsu Hydraulic Oil HO 46-HMDocument5 pages057608komatsu Hydraulic Oil HO 46-HMrolex21No ratings yet
- 601 2023 30Document8 pages601 2023 30Shahram AwanNo ratings yet
- s10787 017 0311 XDocument4 pagess10787 017 0311 X2nmjtzn9rgNo ratings yet
- 13 - BPS Update TOG 2022Document8 pages13 - BPS Update TOG 2022Anna JuniedNo ratings yet
- Gut Microbiota in Neurodegenerative DisordersDocument31 pagesGut Microbiota in Neurodegenerative DisordersMércia FiuzaNo ratings yet
- Isola Etal 2020 Clo IDocument11 pagesIsola Etal 2020 Clo IKAREN ESTHER CAPACOILA GRANADOSNo ratings yet
- Histamine H2 Receptor-Mediated Suppression of Intestinal Inflammation by Probiotic Lactobacillus ReuteriDocument12 pagesHistamine H2 Receptor-Mediated Suppression of Intestinal Inflammation by Probiotic Lactobacillus ReuteriNejc KovačNo ratings yet
- The Role of Prophylactic Ibuprofen and N-Acetylcysteine On The Level of Cytokines in Periapical Exudates and The Post-Treatment PainDocument6 pagesThe Role of Prophylactic Ibuprofen and N-Acetylcysteine On The Level of Cytokines in Periapical Exudates and The Post-Treatment PainNadya PurwantyNo ratings yet
- Article 180803Document2 pagesArticle 180803mustafaali9711No ratings yet
- Review: Mind-Altering With The Gut: Modulation of The Gut-Brain Axis With ProbioticsDocument11 pagesReview: Mind-Altering With The Gut: Modulation of The Gut-Brain Axis With ProbioticsVitor PereiraNo ratings yet
- Fcimb 12 812596Document20 pagesFcimb 12 812596andreafernandez2eNo ratings yet
- Spielman Et Al., 2018-Unhealthy Gut, Unhealthy Brain - The Role of The Intestinal Microbiota in Neurodegenerative DiseasesDocument15 pagesSpielman Et Al., 2018-Unhealthy Gut, Unhealthy Brain - The Role of The Intestinal Microbiota in Neurodegenerative DiseasesEdwar MagallanNo ratings yet
- Journal of Herbal MedicineDocument8 pagesJournal of Herbal MedicineCarlos Andrés Gallardo GarridoNo ratings yet
- Akinyemi Emmanuel ifeoluwa IBD (2)-1-1Document45 pagesAkinyemi Emmanuel ifeoluwa IBD (2)-1-1eakinyemi360No ratings yet
- Mechanisms of Peptic Ulcer Recurrence: Role of in Ammation: ReviewDocument12 pagesMechanisms of Peptic Ulcer Recurrence: Role of in Ammation: ReviewGuilherme Vieira Botelho de Almeida (Teacher Red Balloon Cuiabá)No ratings yet
- The Gut Microbiota in Anxiety and Depression - A Systematic Review (Simpson Et Al, 2020)Document18 pagesThe Gut Microbiota in Anxiety and Depression - A Systematic Review (Simpson Et Al, 2020)GokushimakNo ratings yet
- Akinyemi Emmanuel ifeoluwa IBD (2)-1Document46 pagesAkinyemi Emmanuel ifeoluwa IBD (2)-1eakinyemi360No ratings yet
- 13. MicroRNAs 106b and 222 Improve Hyperglycemia in a Mouse Model of Insulin - Deficient Diabetes via Pancreatic Β-Cell ProliferationDocument10 pages13. MicroRNAs 106b and 222 Improve Hyperglycemia in a Mouse Model of Insulin - Deficient Diabetes via Pancreatic Β-Cell ProliferationvirginiaviolaNo ratings yet
- The_microbiota-gut-brain_axis_regulates_motivationDocument3 pagesThe_microbiota-gut-brain_axis_regulates_motivationchaiyadol tantasithNo ratings yet
- Botulinum Toxin in CancerDocument11 pagesBotulinum Toxin in CancerAlicia Ramirez HernandezNo ratings yet
- Host Modulation Therapy An Updated ReviewDocument4 pagesHost Modulation Therapy An Updated Reviewpaper kitaNo ratings yet
- The Anti Inflammatory Effect of Triphala in Arthritic Induced RatsDocument11 pagesThe Anti Inflammatory Effect of Triphala in Arthritic Induced RatsShyam S SundarNo ratings yet
- Bethany Pinnock - Periodontics V Research Paper (Final)Document5 pagesBethany Pinnock - Periodontics V Research Paper (Final)Bethany PinnockNo ratings yet
- 80 BiotaDocument7 pages80 BiotaAlfonso De la FuenteNo ratings yet
- Raja Et Al 2013Document7 pagesRaja Et Al 2013Sahera Nurhidayah NasutionNo ratings yet
- Potential Influences of Gut Microbiota On The Formation of Intracranial AneurysmDocument14 pagesPotential Influences of Gut Microbiota On The Formation of Intracranial AneurysmMércia FiuzaNo ratings yet
- The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain CommunicationDocument14 pagesThe Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain CommunicationMércia FiuzaNo ratings yet
- Diet MS Valter LongoDocument12 pagesDiet MS Valter LongoAnonymous 0qYv7WpgNo ratings yet
- 1 s2.0 S0163725822002212 MainDocument29 pages1 s2.0 S0163725822002212 MainNélida Lucia Rodriguez AscañoNo ratings yet
- Jurnal SR 3Document12 pagesJurnal SR 3Zaina Khairunisa AsyaNo ratings yet
- Manzhulo 2015Document8 pagesManzhulo 2015Yolla JufandaNo ratings yet
- Animal Models of Rheumatoid Pain: Experimental Systems and InsightsDocument12 pagesAnimal Models of Rheumatoid Pain: Experimental Systems and InsightsLourdes MendietaNo ratings yet
- 10.1515revneuro 2019 0005Document12 pages10.1515revneuro 2019 0005Suhartini RahmanNo ratings yet
- Probiotics Article 2019 PDFDocument10 pagesProbiotics Article 2019 PDFLuiis LimaNo ratings yet
- Share 10.1.1.825.5698Document36 pagesShare 10.1.1.825.5698Aidil CakepNo ratings yet
- Presentation Gut Brain August 2018Document17 pagesPresentation Gut Brain August 2018Sribharathi Indrajith100% (1)
- Tratament Conservator DupDocument3 pagesTratament Conservator DupDora PopaNo ratings yet
- Role of The Gut Microbiota in Ischemic Stroke: Katarzyna Winek, Ulrich Dirnagl, Andreas MeiselDocument7 pagesRole of The Gut Microbiota in Ischemic Stroke: Katarzyna Winek, Ulrich Dirnagl, Andreas MeiselArdianNo ratings yet
- Host Modulation in PeriodonticsDocument12 pagesHost Modulation in PeriodonticsFirma Nurdinia DewiNo ratings yet
- Keaa 192Document11 pagesKeaa 192germanNo ratings yet
- Inflammatory Bowel Disease DayaDocument74 pagesInflammatory Bowel Disease DayarajhiniNo ratings yet
- Role - and - Mechanism - of - Gut MicrobiomaDocument12 pagesRole - and - Mechanism - of - Gut MicrobiomaAyushi MauryaNo ratings yet
- Gut Dysbiosis, Leaky Gut, and Intestinal Epithelial Proliferation in Neurological Disorders Towards The Development of A New Therapeutic Using Amino Acids, Prebiotics, Probiotics, and PostbioticsDocument23 pagesGut Dysbiosis, Leaky Gut, and Intestinal Epithelial Proliferation in Neurological Disorders Towards The Development of A New Therapeutic Using Amino Acids, Prebiotics, Probiotics, and PostbioticsLevente BalázsNo ratings yet
- Nutrients 13 03192 v3Document17 pagesNutrients 13 03192 v3Firman Kusuma AtmajaNo ratings yet
- The Gut Microbiota in Inflammatory Bowel DiseaseDocument14 pagesThe Gut Microbiota in Inflammatory Bowel DiseaseMércia FiuzaNo ratings yet
- Jurnal 3Document20 pagesJurnal 3mardha tillahNo ratings yet
- 2020 - Guo - Oligosaccharadide TMVDocument7 pages2020 - Guo - Oligosaccharadide TMVandreeboneNo ratings yet
- Gastroprotectants in Small Animal Veterinary Practice - A Review of The Evidence Part 1 - Cyto-Protective DrugsDocument16 pagesGastroprotectants in Small Animal Veterinary Practice - A Review of The Evidence Part 1 - Cyto-Protective DrugsPatriciaNo ratings yet
- Molecules 28 00106Document17 pagesMolecules 28 00106Jeoval NetoNo ratings yet
- Lect 1 Host ModulationDocument22 pagesLect 1 Host ModulationrupaliNo ratings yet
- Botox DTM Neuralgia V Par EtcDocument16 pagesBotox DTM Neuralgia V Par EtcjosfranNo ratings yet
- Bromelain A Potential Strategy For The Adjuvant Treatment of PeriodontitisDocument6 pagesBromelain A Potential Strategy For The Adjuvant Treatment of Periodontitismajiamal99No ratings yet
- Article 1 Irfan KhanDocument19 pagesArticle 1 Irfan KhanAshique RajputNo ratings yet
- Format Jurnal Pak Kir 141018Document7 pagesFormat Jurnal Pak Kir 141018daniel_alexander_susenoNo ratings yet
- Cranio®: The Journal of Craniomandibular & Sleep PracticeDocument11 pagesCranio®: The Journal of Craniomandibular & Sleep PracticeAnthony VillafañeNo ratings yet
- Nsaids in PeriodontologyDocument7 pagesNsaids in Periodontologysalman khawarNo ratings yet
- Diacerein New CliicalDocument9 pagesDiacerein New Cliicaledo adimastaNo ratings yet
- Probiotic Antigenotoxic Activity As A DNA Bioprotective Tool: A Minireview With Focus On Endocrine DisruptorsDocument11 pagesProbiotic Antigenotoxic Activity As A DNA Bioprotective Tool: A Minireview With Focus On Endocrine DisruptorsAjit Kumar BaidyaNo ratings yet
- Messengers From The MicrobiotaDocument3 pagesMessengers From The MicrobiotaDra. Ianna CarmoNo ratings yet
- Oral Probiotic Treatment of Lactobacillus Rhamnosus Lcr35® Prevents Visceral Hypersensitivity To A Colonic Inflammation and An Acute Psychological StressDocument13 pagesOral Probiotic Treatment of Lactobacillus Rhamnosus Lcr35® Prevents Visceral Hypersensitivity To A Colonic Inflammation and An Acute Psychological StressMaria TejedaNo ratings yet
- The new frontier of anti-aging medicine: Miracles nutrigenomics senolitic products for a healthy lifeFrom EverandThe new frontier of anti-aging medicine: Miracles nutrigenomics senolitic products for a healthy lifeNo ratings yet
- jcdr-9-ZE08Document4 pagesjcdr-9-ZE08germanNo ratings yet
- The Use of Pharmacologic Agents in The Management of Temporomandibular Joint DisorderDocument7 pagesThe Use of Pharmacologic Agents in The Management of Temporomandibular Joint DisordergermanNo ratings yet
- The Problems Preventing Progress in Adhesive Restorative DentistryDocument6 pagesThe Problems Preventing Progress in Adhesive Restorative DentistrygermanNo ratings yet
- Ribeiro 2020Document8 pagesRibeiro 2020germanNo ratings yet
- Keaa 192Document11 pagesKeaa 192germanNo ratings yet
- Meta 2.1 Ocampo Libro (15-21)Document7 pagesMeta 2.1 Ocampo Libro (15-21)germanNo ratings yet
- Meta 2.1 Ocampo LibroDocument21 pagesMeta 2.1 Ocampo LibrogermanNo ratings yet
- Meta 2.1 Ocampo Libro (08-14)Document7 pagesMeta 2.1 Ocampo Libro (08-14)germanNo ratings yet
- Meta 2.1 Ocampo Libro (01-07)Document7 pagesMeta 2.1 Ocampo Libro (01-07)germanNo ratings yet
- Meta 1.1 Ocampo Libro (11-21)Document11 pagesMeta 1.1 Ocampo Libro (11-21)germanNo ratings yet
- Meta 1.1 Ocampo Libro (11-21) (01-06)Document6 pagesMeta 1.1 Ocampo Libro (11-21) (01-06)germanNo ratings yet
- Meta 2.2 Ocampo LibroDocument7 pagesMeta 2.2 Ocampo LibrogermanNo ratings yet
- "After All, There Is Nothing As Interesting As People, and One Can Never Study Them Enough" Vincent Van GoghDocument38 pages"After All, There Is Nothing As Interesting As People, and One Can Never Study Them Enough" Vincent Van GoghAnonymous 1gH7ra9ANo ratings yet
- Hyperworks Technology Conference (HTC) 2011: Optimization of PMH StructuresDocument25 pagesHyperworks Technology Conference (HTC) 2011: Optimization of PMH StructuresgiakovidNo ratings yet
- Stresses and Challenges For New Graduates in HospitalsDocument6 pagesStresses and Challenges For New Graduates in HospitalskitsilNo ratings yet
- Omonigbehin Emmanuel AdedayoDocument12 pagesOmonigbehin Emmanuel AdedayoAriel GhieNo ratings yet
- 5X5 Intermediate - Bill StarrDocument7 pages5X5 Intermediate - Bill StarrAfrican MastiffNo ratings yet
- Which of The Following Is NOT True About Dust?Document3 pagesWhich of The Following Is NOT True About Dust?lucel palaca100% (1)
- Coriander (Corindrum Sativum L) and Its Bioactive ConstituentsDocument19 pagesCoriander (Corindrum Sativum L) and Its Bioactive ConstituentsFirda AuliyaNo ratings yet
- 10besar Kode PenyakitDocument5 pages10besar Kode PenyakitDEWINo ratings yet
- Closed PTW AuditDocument1 pageClosed PTW Auditf.BNo ratings yet
- Benefits of Hypnosis Author Mental Health America of Northern KentuckyDocument4 pagesBenefits of Hypnosis Author Mental Health America of Northern KentuckyJakarta SelamatNo ratings yet
- Health Quiz No. 2: NAME: - GRADE 6 - SCOREDocument2 pagesHealth Quiz No. 2: NAME: - GRADE 6 - SCORELove LeiNo ratings yet
- Hope - 1 Grade 11: Exercise For FitnessDocument6 pagesHope - 1 Grade 11: Exercise For FitnessTABBAL JAN PAOLONo ratings yet
- Margaret Paul - Taking The Lid Off Your FeelingsDocument4 pagesMargaret Paul - Taking The Lid Off Your FeelingspapetologeneNo ratings yet
- Medical Condition Report Free PDF TemplateDocument2 pagesMedical Condition Report Free PDF Templateapi-340236228No ratings yet
- WEEK 2: 12A, 12B, 12G Ôn Tập Khảo Sát Pronunciation ClosestDocument6 pagesWEEK 2: 12A, 12B, 12G Ôn Tập Khảo Sát Pronunciation ClosestHàNhậtNguyễnNo ratings yet
- Development of Push-Pull Osmotic Pump Tablets For A SlightlyDocument4 pagesDevelopment of Push-Pull Osmotic Pump Tablets For A SlightlyphamuyenthuNo ratings yet
- Life Calling Map Fillable FormDocument2 pagesLife Calling Map Fillable Formapi-300853489No ratings yet
- The Effects of Emotion On Short-Term Power Spectrum Analysis of Heart RateDocument5 pagesThe Effects of Emotion On Short-Term Power Spectrum Analysis of Heart RateManuel Guerrero GómezNo ratings yet
- Anna Garcia Death Timeline 1Document19 pagesAnna Garcia Death Timeline 1api-245176779No ratings yet
- Rose Doe v. The City of New York, Et. Al.Document57 pagesRose Doe v. The City of New York, Et. Al.Daily Caller News FoundationNo ratings yet
- Journal Medicine: The New EnglandDocument7 pagesJournal Medicine: The New EnglandTony RamirezNo ratings yet
- Knowledge ManagementDocument63 pagesKnowledge Managementrahul-singh-6592No ratings yet
- National Program For Rehabilitation of Polluted Site - A Case StudyDocument4 pagesNational Program For Rehabilitation of Polluted Site - A Case StudyijeteeditorNo ratings yet
- PREGNANCYDocument4 pagesPREGNANCYShaileeNo ratings yet
- Daftar Obat %pareto Dan Ven 15Document200 pagesDaftar Obat %pareto Dan Ven 15sofia nofiantiNo ratings yet
- UP Vaccine Center ListDocument106 pagesUP Vaccine Center ListRamendra Nath JhaNo ratings yet
- Language Disorders - 5Document27 pagesLanguage Disorders - 5Marta Sampedro GonzalezNo ratings yet
- Henares Vs LTFRBDocument2 pagesHenares Vs LTFRBCarolina Lim GambanNo ratings yet
- Maternity Nursing ReviewDocument37 pagesMaternity Nursing Reviewshenric16No ratings yet