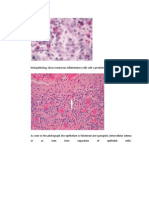
Download as pdf or txt
You might also like
- AACR 2022 Proceedings: Part A Online-Only and April 10From EverandAACR 2022 Proceedings: Part A Online-Only and April 10No ratings yet
- Path DumpsDocument44 pagesPath DumpsAndleeb ImranNo ratings yet
- Ijms 24 06174Document17 pagesIjms 24 06174Naga MuthuNo ratings yet
- Role of Galectins in Inflammatory and Immunomodulatory Processes 2002Document11 pagesRole of Galectins in Inflammatory and Immunomodulatory Processes 2002evilbioNo ratings yet
- CSF3R 6Document7 pagesCSF3R 6AndikhaNo ratings yet
- 1 s2.0 S0022282815001583 MainDocument13 pages1 s2.0 S0022282815001583 MainRicardo GomezNo ratings yet
- TS Glycobiology in Cancer EbookDocument8 pagesTS Glycobiology in Cancer EbookDaisy Joanna Castañeda MataNo ratings yet
- Prognostic Factors For Chronic Lymphocytic LeukemiaDocument6 pagesPrognostic Factors For Chronic Lymphocytic LeukemiaJose AbadiaNo ratings yet
- Impact of Dietary Components On NK and Treg Cell Function For Cancer PreventionDocument6 pagesImpact of Dietary Components On NK and Treg Cell Function For Cancer PreventionEgy SunandaNo ratings yet
- Integrins As Therapeutic TargetsDocument8 pagesIntegrins As Therapeutic TargetsAnonymous ceYk4p4No ratings yet
- Sobrecarga BogotaDocument11 pagesSobrecarga BogotaAndrés Felipe PérezNo ratings yet
- The oncometabolite 2-hydroxyglutarate inhibits microglial activation via the AMPK/mTOR/NF-κB pathwayDocument11 pagesThe oncometabolite 2-hydroxyglutarate inhibits microglial activation via the AMPK/mTOR/NF-κB pathwayPaige MunroeNo ratings yet
- KDM5B MacrophageDocument14 pagesKDM5B MacrophageMaurício DallowayNo ratings yet
- Cancer Stem Cells: Potential Target For Bioactive Food ComponentsDocument8 pagesCancer Stem Cells: Potential Target For Bioactive Food ComponentsMikhail PisarevNo ratings yet
- Immune Cells in Liver RegenerationDocument12 pagesImmune Cells in Liver RegenerationJesus_Arraiz_14No ratings yet
- TS Glycobiology in Cancer EbookDocument8 pagesTS Glycobiology in Cancer EbookDaisy Joanna Castañeda MataNo ratings yet
- Jak Stat InhibitorsDocument64 pagesJak Stat InhibitorsSupratim KarmakarNo ratings yet
- Jcem 1404Document8 pagesJcem 1404lucylit0666No ratings yet
- Therapeutic Potential of Galectin-3 Inhibition - Slack 2021Document12 pagesTherapeutic Potential of Galectin-3 Inhibition - Slack 2021Stuart Fysh100% (1)
- Leuemia MarkerDocument14 pagesLeuemia MarkerAdna SantanaNo ratings yet
- Mast Cell Activation DiseaseDocument9 pagesMast Cell Activation DiseaseKata TölgyesiNo ratings yet
- Gaucher Disease - Pathogenesis, Clinical Manifestations, and Diagnosis - UpToDateDocument33 pagesGaucher Disease - Pathogenesis, Clinical Manifestations, and Diagnosis - UpToDateAnna MoraNo ratings yet
- At The Crossroads of Lipid Metabolism and InflammationDocument7 pagesAt The Crossroads of Lipid Metabolism and InflammationChow Sing SingNo ratings yet
- Expresion Patterns of Genes, Regulating Lipid Metabolism in Prostate TumorsDocument16 pagesExpresion Patterns of Genes, Regulating Lipid Metabolism in Prostate TumorsАнна ШаповаловаNo ratings yet
- Exploratory Assessment of Galectin-1 - 3 and - 9 inDocument17 pagesExploratory Assessment of Galectin-1 - 3 and - 9 inJCNo ratings yet
- HHS Public AccessDocument23 pagesHHS Public AccessshazaNo ratings yet
- El Glicocalix y La SepsisDocument12 pagesEl Glicocalix y La SepsisHector Carlos Camacho AguirreNo ratings yet
- Mendel Et Al 2003Document12 pagesMendel Et Al 2003Selliana Maretha Wijaya KusumaNo ratings yet
- NIH Public Access: Author ManuscriptDocument15 pagesNIH Public Access: Author ManuscriptarthoclaseNo ratings yet
- Abedalthagaf 2018Document8 pagesAbedalthagaf 2018NicoNo ratings yet
- Rodrguez Vicente 2013Document14 pagesRodrguez Vicente 2013Jose AbadiaNo ratings yet
- Downregulation of Dystroglycan Glycosyltransferases Large2 and Ispd Associate With Increased Mortality in Clear Cell Renal Cell CarcinomaDocument12 pagesDownregulation of Dystroglycan Glycosyltransferases Large2 and Ispd Associate With Increased Mortality in Clear Cell Renal Cell CarcinomaAlex HydronNo ratings yet
- Galectin-3 Is Independently Associated With Progression of Nephropathy in Type 2 Diabetes MellitusDocument8 pagesGalectin-3 Is Independently Associated With Progression of Nephropathy in Type 2 Diabetes Mellituslucylit0666No ratings yet
- Jurnal MG MyastheniaDocument11 pagesJurnal MG MyastheniaTomy NugrohoNo ratings yet
- 2019 Vaamonde-GarciaDocument13 pages2019 Vaamonde-GarciaHistologia elementarNo ratings yet
- Epidemiology, Pathogenesis, and Clinical Manifestations of Celiac Disease in Adults - UpToDateDocument41 pagesEpidemiology, Pathogenesis, and Clinical Manifestations of Celiac Disease in Adults - UpToDatealinetibes1No ratings yet
- Key WordsDocument8 pagesKey WordsThanasis PapatheodorouNo ratings yet
- FCVM 09 868372Document14 pagesFCVM 09 868372lucylit0666No ratings yet
- Journal Club Terapia Con Células T Del Receptor de Antígeno Quimérico anti-CD19 para El Lupus Eritematoso Sistémico RefractarioDocument5 pagesJournal Club Terapia Con Células T Del Receptor de Antígeno Quimérico anti-CD19 para El Lupus Eritematoso Sistémico RefractarioCristhian RodriguezNo ratings yet
- FOXP3 Variants Are Independently Associated With Transforming Growth Factor B1 Plasma Levels in Female Patients With Inflammatory Bowel DiseaseDocument9 pagesFOXP3 Variants Are Independently Associated With Transforming Growth Factor B1 Plasma Levels in Female Patients With Inflammatory Bowel DiseasePaula KikuchiNo ratings yet
- Normal Value CytokinesDocument7 pagesNormal Value Cytokinesdiana.alyNo ratings yet
- Some Serum Cytokines in Celiac Disease, Non-CeliacDocument10 pagesSome Serum Cytokines in Celiac Disease, Non-CeliacCentral Asian StudiesNo ratings yet
- HHS Public Access: Studying Polyglutamine Diseases in DrosophilaDocument39 pagesHHS Public Access: Studying Polyglutamine Diseases in DrosophilaJoeNo ratings yet
- 13GLUT1 overexpression enhances CAR T cellDocument13 pages13GLUT1 overexpression enhances CAR T cellzjq1432031523No ratings yet
- Galectins As Modulators of Tumour Progression: Fu-Tong Liu and Gabriel A. RabinovichDocument13 pagesGalectins As Modulators of Tumour Progression: Fu-Tong Liu and Gabriel A. RabinovichjmbattagliottiNo ratings yet
- Royal JellyDocument10 pagesRoyal JellyninukNo ratings yet
- Cytoglobin Inhibits Migration Through PI3K/AKT/mTOR Pathway in Fibroblast CellsDocument10 pagesCytoglobin Inhibits Migration Through PI3K/AKT/mTOR Pathway in Fibroblast CellsDilay CelikNo ratings yet
- Anti-Proliferative Properties of Thymoquinone To Overcome Cisplatin-Induced Drug Resistance in SKOV-3Document8 pagesAnti-Proliferative Properties of Thymoquinone To Overcome Cisplatin-Induced Drug Resistance in SKOV-3mehulshah1126No ratings yet
- Ijms 24 09850Document26 pagesIjms 24 09850Damiana MargineanuNo ratings yet
- University of Medical Sciences and Technology Faculty of PharmacyDocument7 pagesUniversity of Medical Sciences and Technology Faculty of PharmacyTalal Ahmed Awad MohammedNo ratings yet
- 32.mast Cell Silencing A Novel Therapeutic Approach For Urticaria and Other Mast Cell MediatedDocument15 pages32.mast Cell Silencing A Novel Therapeutic Approach For Urticaria and Other Mast Cell MediatedBasil KNo ratings yet
- DMMMDocument8 pagesDMMMvivitanovNo ratings yet
- Glicocorticoide e MineralizaçãoDocument9 pagesGlicocorticoide e Mineralizaçãomatheus galvãoNo ratings yet
- C2C12 Weiwei Chu 2016Document7 pagesC2C12 Weiwei Chu 2016Fujiko Saavedra LeivaNo ratings yet
- CC 11 3611Document16 pagesCC 11 3611Sergeat18BNo ratings yet
- Obesidad y LeucemiaDocument5 pagesObesidad y LeucemiaAlejandro PalayNo ratings yet
- GADD45B Growth Arrest and DNA-damage-inducible, BetaDocument6 pagesGADD45B Growth Arrest and DNA-damage-inducible, BetaHassan Ahmed HassanNo ratings yet
- Basophil Activation Test: Old and New Applications in AllergyDocument12 pagesBasophil Activation Test: Old and New Applications in AllergyAybars DenizNo ratings yet
- Ajtr 0151665Document12 pagesAjtr 0151665Muhammad ImranNo ratings yet
- JAK3 InhibitorDocument13 pagesJAK3 Inhibitorsumera120488No ratings yet
- Gastric Cancer GlycosilationDocument13 pagesGastric Cancer GlycosilationJosué VelázquezNo ratings yet
- Protective Dural Flap For Bone Drilling at The Paraclinoid Region and Porus AcusticusDocument3 pagesProtective Dural Flap For Bone Drilling at The Paraclinoid Region and Porus AcusticusVenansius Ratno KurniawanNo ratings yet
- Role of Transcranial Approaches in The Treatment of Sellar and Suprasellar LesionsDocument28 pagesRole of Transcranial Approaches in The Treatment of Sellar and Suprasellar LesionsVenansius Ratno Kurniawan100% (1)
- Immersive Surgical Anatomy of The Pterional ApproachDocument25 pagesImmersive Surgical Anatomy of The Pterional ApproachVenansius Ratno KurniawanNo ratings yet
- Altitude Illness - Risk Factors, Prevention, Presentation, and TreatmentDocument8 pagesAltitude Illness - Risk Factors, Prevention, Presentation, and TreatmentVenansius Ratno KurniawanNo ratings yet
- Cardiovascular Parameter in A Mixed-Sex Swine Study of Severe Dcs Sickness Treated With The Emulsified Perfluorocarbon OxycyteDocument9 pagesCardiovascular Parameter in A Mixed-Sex Swine Study of Severe Dcs Sickness Treated With The Emulsified Perfluorocarbon OxycyteVenansius Ratno KurniawanNo ratings yet
- Decompression Sickness Treatment: First AidDocument8 pagesDecompression Sickness Treatment: First AidVenansius Ratno KurniawanNo ratings yet
- AKI Due To DCSDocument3 pagesAKI Due To DCSVenansius Ratno KurniawanNo ratings yet
- Update ACLSDocument64 pagesUpdate ACLSVenansius Ratno KurniawanNo ratings yet
- Immunity and Immunopathology To Viruses - What Decides The Outcome?Document23 pagesImmunity and Immunopathology To Viruses - What Decides The Outcome?Venansius Ratno KurniawanNo ratings yet
- Expression of Fibroblast Growth Factor 23, Vitamin D Receptor, and Sclerostin in Bone Tissue From Hypercalciuric Stone FormersDocument8 pagesExpression of Fibroblast Growth Factor 23, Vitamin D Receptor, and Sclerostin in Bone Tissue From Hypercalciuric Stone FormersVenansius Ratno KurniawanNo ratings yet
- Pathopysiology of AdenomyosisDocument11 pagesPathopysiology of AdenomyosisVenansius Ratno KurniawanNo ratings yet
- Gambar Conjungtivitis VernalDocument6 pagesGambar Conjungtivitis VernalVenansius Ratno KurniawanNo ratings yet
- Cellular Aberrations (Basic Concepts of Oncology)Document12 pagesCellular Aberrations (Basic Concepts of Oncology)PATRIZJA YSABEL REYESNo ratings yet
- Oral Pathology & Oral Microbiology: SyllabusDocument14 pagesOral Pathology & Oral Microbiology: SyllabusElijah JoeNo ratings yet
- Odontomes and Odontogenic TumoursDocument16 pagesOdontomes and Odontogenic TumourssarahNo ratings yet
- Histology and HistopathologyDocument42 pagesHistology and HistopathologyDreyden HaloNo ratings yet
- Class 6 PathologyDocument4 pagesClass 6 PathologyMostafizur RahmanNo ratings yet
- What Is Cancer? What Causes Cancer?: Discovered An Important Clue As To Why Cancer Cells SpreadDocument8 pagesWhat Is Cancer? What Causes Cancer?: Discovered An Important Clue As To Why Cancer Cells SpreadAlfred Melvin SolivaNo ratings yet
- Brain Tumor Segmentation and Detection Using Nueral NetworksDocument9 pagesBrain Tumor Segmentation and Detection Using Nueral Networksjoshi manoharNo ratings yet
- Brochure Services IHC StainsDocument16 pagesBrochure Services IHC StainsYousra ZeidanNo ratings yet
- Instructor's Presentation-Tumor MarkersDocument57 pagesInstructor's Presentation-Tumor MarkersMACOB, ETHELHYN JHANENo ratings yet
- Radiologic Pathology Intro 1Document50 pagesRadiologic Pathology Intro 1James MoralesNo ratings yet
- Pinto 2012A fin de cuantificar la asociación entre la exposición de mujeres gestantes a una serie de supuestos factores de riesgo y la presencia de bajo peso al nacer (BPN) en sus productos, entre mayo y septiembre de 1996 se realizó un estudio epidemiológico en la Provincia N de un país de América del Sur. Se definió como enfermo a toda madre que haya parido un niño/a con BPN (menos de 2.500 gramos) y como no enfermo a toda madre que haya parido un niño/a sin BPN. Se seleccionaron 1.556 enfermos y 16.910 no enfermos, registrados entre 1988 y 1995 en la base de datos del sistema informático perinatal disponible en dicha provincia. El Cuadro 3.18 presenta un extracto de los resultados de dicho estudio.Document26 pagesPinto 2012A fin de cuantificar la asociación entre la exposición de mujeres gestantes a una serie de supuestos factores de riesgo y la presencia de bajo peso al nacer (BPN) en sus productos, entre mayo y septiembre de 1996 se realizó un estudio epidemiológico en la Provincia N de un país de América del Sur. Se definió como enfermo a toda madre que haya parido un niño/a con BPN (menos de 2.500 gramos) y como no enfermo a toda madre que haya parido un niño/a sin BPN. Se seleccionaron 1.556 enfermos y 16.910 no enfermos, registrados entre 1988 y 1995 en la base de datos del sistema informático perinatal disponible en dicha provincia. El Cuadro 3.18 presenta un extracto de los resultados de dicho estudio.andresNo ratings yet
- Pathology ofDocument114 pagesPathology ofRahul Audenesen BratNo ratings yet
- Chapter 21 - The Lower Urinary Tract and Male Genital SystemDocument38 pagesChapter 21 - The Lower Urinary Tract and Male Genital SystemAgnieszka WisniewskaNo ratings yet
- Adenoma Carcinoma SequenceDocument27 pagesAdenoma Carcinoma SequenceKhurshedul AlamNo ratings yet
- PDF Ferris Clinical Advisor 2019 5 Books in 1 Fred F Ferri Ebook Full ChapterDocument53 pagesPDF Ferris Clinical Advisor 2019 5 Books in 1 Fred F Ferri Ebook Full Chapterherman.palmo638100% (3)
- Current Concepts in Plastic Surgery - AgulloDocument273 pagesCurrent Concepts in Plastic Surgery - AgulloAnonymous LnWIBo1GNo ratings yet
- UntitledDocument3 pagesUntitled佳昇.No ratings yet
- Uk Guidelines SarcomaDocument21 pagesUk Guidelines Sarcomachu_chiang_3No ratings yet
- NeoplasiaDocument144 pagesNeoplasiaAsa DullahNo ratings yet
- Pathology Lecture SeriesDocument168 pagesPathology Lecture SeriesButch DumdumNo ratings yet
- Plac Pathology - Drucilla RobertsDocument11 pagesPlac Pathology - Drucilla RobertsconstantinilieNo ratings yet
- Pigmented Oral LesionDocument133 pagesPigmented Oral LesionArnela UniqueNo ratings yet
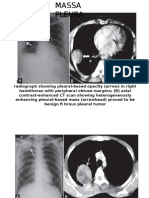
- Massa PleuraDocument4 pagesMassa PleuraPrayogi KramyNo ratings yet
- Tissue MicroarrayDocument22 pagesTissue MicroarrayNithin DiwagarNo ratings yet
- Cancer Mother of All Diseases ASU Study Guide Exam #1Document2 pagesCancer Mother of All Diseases ASU Study Guide Exam #1Nitin NandaNo ratings yet
- Health Insurance - 202316222434Document54 pagesHealth Insurance - 202316222434AayushNo ratings yet
- HarshMohan PDFDocument634 pagesHarshMohan PDFjothika50% (2)
- What Are The Biochemical and Structural Changes That Occur in Cancer CellDocument7 pagesWhat Are The Biochemical and Structural Changes That Occur in Cancer CellsujsamNo ratings yet
- Urinary Bladder Diverticulum and Its Association With Malignancy: An Anatomical Study On CadaversDocument3 pagesUrinary Bladder Diverticulum and Its Association With Malignancy: An Anatomical Study On CadaversDwi Tantri SPNo ratings yet