
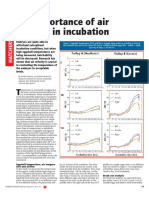
Modeling Incubation Temperature: The Effects of Incubator Design, Embryonic Development, and Egg Size
Modeling Incubation Temperature: The Effects of Incubator Design, Embryonic Development, and Egg Size
You might also like
- ENEN-635 Environmental Modeling Lab-3 Report - Rev1Document10 pagesENEN-635 Environmental Modeling Lab-3 Report - Rev1Muhammad NaeemNo ratings yet
- Temperature Distribution PDFDocument5 pagesTemperature Distribution PDFEstefania ArizagaNo ratings yet
- 1 s2.0 S0032579119445367 MainDocument9 pages1 s2.0 S0032579119445367 Mainjimmy kNo ratings yet
- The Critical Importance of Incubation Temperature: Nick A. FrenchDocument5 pagesThe Critical Importance of Incubation Temperature: Nick A. FrenchThE BoNg TeChniCaLNo ratings yet
- Lin 2005Document7 pagesLin 2005Jake Floyd G. FabianNo ratings yet
- Heat and Moisture Production of Broilers Kept On Straw BeddingDocument11 pagesHeat and Moisture Production of Broilers Kept On Straw BeddingAlejandro OviedoNo ratings yet
- Body Temperatures in Free - Ying PigeonsDocument5 pagesBody Temperatures in Free - Ying PigeonsDummy DonNo ratings yet
- Chick Quality2Document12 pagesChick Quality2Dani GarnidaNo ratings yet
- Effect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualityDocument6 pagesEffect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualitySebastian RodriguezNo ratings yet
- (Apis Mellifera) Foraging in The Sonoran Desert: Temperature Regulation of Honey BeesDocument16 pages(Apis Mellifera) Foraging in The Sonoran Desert: Temperature Regulation of Honey BeeskarlaNo ratings yet
- Arch Dis Child 1975 Aynsley Green 215 9Document6 pagesArch Dis Child 1975 Aynsley Green 215 9syncmaster1234567890No ratings yet
- Thermoregulation in PoultryDocument7 pagesThermoregulation in PoultryDilip GuptaNo ratings yet
- Erjsp.2011.51.56 Vol ConsiderationDocument6 pagesErjsp.2011.51.56 Vol Considerationoluchukwu u.No ratings yet
- 1 s2.0 S0032579119439527 MainDocument11 pages1 s2.0 S0032579119439527 Mainkumpul tugasNo ratings yet
- Conditions For Cold Stress Development in Dairy Cattle Kept in Free Stall Barn During Severe FrostsDocument8 pagesConditions For Cold Stress Development in Dairy Cattle Kept in Free Stall Barn During Severe FrostsMuhammad Firdaus FebriansyahNo ratings yet
- A Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsDocument8 pagesA Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsSebastian RodriguezNo ratings yet
- Seasonal Temperature Effects and Predicting Development Rates of Marine Copepod EggsDocument7 pagesSeasonal Temperature Effects and Predicting Development Rates of Marine Copepod EggsJordan IsmaelNo ratings yet
- Wasmen HEAT STRESSDocument4 pagesWasmen HEAT STRESSByki HaqiNo ratings yet
- Modelling of Heat and Mass Transfer Processes in NeonatologyDocument12 pagesModelling of Heat and Mass Transfer Processes in NeonatologySai Santhosh ManepallyNo ratings yet
- Biocarbon Production From Hungarian Sunflower ShellsDocument5 pagesBiocarbon Production From Hungarian Sunflower ShellsMuhamad SuharNo ratings yet
- 2009.10.13 Design and Performance Evaluation of Automatic Egg IncubatorDocument13 pages2009.10.13 Design and Performance Evaluation of Automatic Egg IncubatorJohn Taulo100% (1)
- Regulacion de Temperatura AvesDocument32 pagesRegulacion de Temperatura AvesSam Lopez LopezNo ratings yet
- 08a8 PDFDocument18 pages08a8 PDFstteevennnNo ratings yet
- 1 s2.0 S0022474X14000435 Main PDFDocument6 pages1 s2.0 S0022474X14000435 Main PDFMario CalderonNo ratings yet
- Journal of Stored Products Research: R. Rusinek, R. Koby 1kaDocument6 pagesJournal of Stored Products Research: R. Rusinek, R. Koby 1kaMario CalderonNo ratings yet
- Brood Production Increases When Artificial Heating Is Provided To Colonies of Stingless BeesDocument6 pagesBrood Production Increases When Artificial Heating Is Provided To Colonies of Stingless BeesOtaviano Carneiro Da Cunha NetoNo ratings yet
- Group-2 ChE-135 Post Lab Experiment-1Document18 pagesGroup-2 ChE-135 Post Lab Experiment-1Rievelyn MartusNo ratings yet
- Heat Transfer Process TUT (After Mid Semester)Document11 pagesHeat Transfer Process TUT (After Mid Semester)vaishnavi singh100% (1)
- Susceptibility of Broiler Chickens To Hemorrhages in Muscles: The Effect of Stock and Rearing Temperature RegimenDocument8 pagesSusceptibility of Broiler Chickens To Hemorrhages in Muscles: The Effect of Stock and Rearing Temperature RegimenEbrahim AbdullaNo ratings yet
- Analisis Dan Simulasi Distribusi Suhu Udara Pada Kandang Sapi Perah Menggunakan Computational Fluid Dynamics (CFD)Document11 pagesAnalisis Dan Simulasi Distribusi Suhu Udara Pada Kandang Sapi Perah Menggunakan Computational Fluid Dynamics (CFD)SatriantyNo ratings yet
- Specific Heat of Agricultural and Food MaterialsDocument29 pagesSpecific Heat of Agricultural and Food MaterialskqNo ratings yet
- Lumped Heat Transfer AnalysisDocument29 pagesLumped Heat Transfer AnalysisprasannaNo ratings yet
- Temperature and Heat 3Document18 pagesTemperature and Heat 3Samud MorrisNo ratings yet
- Honeybee Colony Thermoregulation - Regulatory Mechanisms and Contribution of Individuals in Dependence On Age, Location and Thermal StressDocument13 pagesHoneybee Colony Thermoregulation - Regulatory Mechanisms and Contribution of Individuals in Dependence On Age, Location and Thermal Stresslosaron437No ratings yet
- Beehive - Temperature and Humidity ControlDocument7 pagesBeehive - Temperature and Humidity ControlJoao FerreiraNo ratings yet
- Thermal Properties of Butter: Peratures Between 17 and - 35°CDocument4 pagesThermal Properties of Butter: Peratures Between 17 and - 35°CpeshoaNo ratings yet
- Investigation of Effective Thermal Conductivity Kinetics of Crust and Core Regions of Potato During Deep Fat Frying JFEDocument6 pagesInvestigation of Effective Thermal Conductivity Kinetics of Crust and Core Regions of Potato During Deep Fat Frying JFELucas PanNo ratings yet
- Acclimation To Hypothermic Incubation in Developing Chicken Embryos (Gallus Domesticus)Document10 pagesAcclimation To Hypothermic Incubation in Developing Chicken Embryos (Gallus Domesticus)Sharon TarquiniNo ratings yet
- TMP 56 BDDocument4 pagesTMP 56 BDFrontiersNo ratings yet
- Calulation of Heat Sterilization Time of Fermentation MediaDocument8 pagesCalulation of Heat Sterilization Time of Fermentation Mediahansenmike698105No ratings yet
- Kustermann1982 RespirationDocument12 pagesKustermann1982 RespirationLiên Nguyễn ThùyNo ratings yet
- ETUDE - Energic Optimisation of Foraging Honeybees - Flexible Change of Strategies in Response To Environmental ChallengesDocument8 pagesETUDE - Energic Optimisation of Foraging Honeybees - Flexible Change of Strategies in Response To Environmental Challengeslosaron437No ratings yet
- ETUDE - Contribution of Honeybee Drones of Different Age To ColonialDocument17 pagesETUDE - Contribution of Honeybee Drones of Different Age To Coloniallosaron437No ratings yet
- INTRODUCTIONDocument3 pagesINTRODUCTIONLeonessa FlorNo ratings yet
- 2 IpekDocument4 pages2 IpekDani GarnidaNo ratings yet
- Untitled DocumentDocument6 pagesUntitled DocumentmandyngoaNo ratings yet
- Heat Stress in AnimalsDocument6 pagesHeat Stress in AnimalsAnke NemirovskyNo ratings yet
- Nordica RocketDocument14 pagesNordica RocketradianNo ratings yet
- Tugas Ehan Perpindahan PanasDocument44 pagesTugas Ehan Perpindahan PanasEHANNo ratings yet
- CAT 2019 Slot 1 RC 2 How Emperor Penguins Survive Antarctica's ChillDocument4 pagesCAT 2019 Slot 1 RC 2 How Emperor Penguins Survive Antarctica's ChillMana GargiNo ratings yet
- Reanimation of Rats From Body Temperatures Between 0 and 1°c by Microwave DiathermyDocument6 pagesReanimation of Rats From Body Temperatures Between 0 and 1°c by Microwave DiathermyLJNo ratings yet
- Prediction of Chilling Times of Foods in Situations Where Evaporative Cooling Is Significant-Part 1. Method DevelopmentDocument15 pagesPrediction of Chilling Times of Foods in Situations Where Evaporative Cooling Is Significant-Part 1. Method DevelopmentMaurício de OliveiraNo ratings yet
- 2022 Computers (Artigo Dissertação)Document10 pages2022 Computers (Artigo Dissertação)marcelo.bahuti2No ratings yet
- 13 Space Heating and Cooling: Properties, Production, ApplicationsDocument1 page13 Space Heating and Cooling: Properties, Production, ApplicationsPon lathaNo ratings yet
- Tazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRDocument6 pagesTazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRSebastian RodriguezNo ratings yet
- ChE 0101 Final ReportDocument29 pagesChE 0101 Final ReportPaartiban PaneerselvamNo ratings yet
- Rep-Jrf 114 2 001Document16 pagesRep-Jrf 114 2 001siyopin173No ratings yet
- The Importance of Air Velocity in IncubationDocument2 pagesThe Importance of Air Velocity in Incubationamamùra maamarNo ratings yet
- Inspires Gas Temperature Un NeonatesDocument5 pagesInspires Gas Temperature Un NeonatesGuillermo MartínezNo ratings yet
- Study of The Bread Baking Process - II. Mathematical ModellingtDocument16 pagesStudy of The Bread Baking Process - II. Mathematical Modellingtmatias andradeNo ratings yet
- Heat and Moisture Transfer between Human Body and EnvironmentFrom EverandHeat and Moisture Transfer between Human Body and EnvironmentNo ratings yet
- Robot RaplimDocument1 pageRobot RaplimSebastian Rodriguez100% (1)
- Diagrama2 PDFDocument1 pageDiagrama2 PDFSebastian RodriguezNo ratings yet
- Food Control: A A B C BDocument9 pagesFood Control: A A B C BSebastian RodriguezNo ratings yet
- Diagrama4 PDFDocument1 pageDiagrama4 PDFSebastian RodriguezNo ratings yet
- Experimental Study of The Dependence of Embryonic Development of Trachurus Trachurus Eggs On TemperatureDocument8 pagesExperimental Study of The Dependence of Embryonic Development of Trachurus Trachurus Eggs On TemperatureSebastian RodriguezNo ratings yet
- Tazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRDocument6 pagesTazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRSebastian RodriguezNo ratings yet
- Using Multinomial Models To Analyse Data From Iberian Sardine Egg Incubation Experiments: A Comparison With Traditional TechniquesDocument9 pagesUsing Multinomial Models To Analyse Data From Iberian Sardine Egg Incubation Experiments: A Comparison With Traditional TechniquesSebastian RodriguezNo ratings yet
- Effect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualityDocument6 pagesEffect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualitySebastian RodriguezNo ratings yet
- A Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsDocument8 pagesA Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsSebastian RodriguezNo ratings yet
- Module 5 - Newtons LawsDocument11 pagesModule 5 - Newtons LawsKrizel CalasaraNo ratings yet
- 3.3a VSEPR Theory PDFDocument21 pages3.3a VSEPR Theory PDFSarvesh Kanaujia100% (8)
- Specular Reflection FTIR: A Non-Contact Method For Analyzing Coatings On Photographs and Other Cultural MaterialsDocument15 pagesSpecular Reflection FTIR: A Non-Contact Method For Analyzing Coatings On Photographs and Other Cultural MaterialsAndres Camilo Gonzalez RomeroNo ratings yet
- Good..... Analysis and Controller Design For Standalone VSIs in Synchronous Reference FrameDocument10 pagesGood..... Analysis and Controller Design For Standalone VSIs in Synchronous Reference FrameAQEEL ANWARNo ratings yet
- Arduino CertificationDocument8 pagesArduino Certificationhack reportNo ratings yet
- Practive Problems Prepared By: Engr. Dean Domenique E. ClidoroDocument11 pagesPractive Problems Prepared By: Engr. Dean Domenique E. ClidoroGrAxieQuidOrNo ratings yet
- Airgun ArraysDocument21 pagesAirgun ArraysGianni PavanNo ratings yet
- Piperazine Impregnation On Zeolite 13X As A Novel Adsorbent For CO Capture: Experimental and ModelingDocument18 pagesPiperazine Impregnation On Zeolite 13X As A Novel Adsorbent For CO Capture: Experimental and ModelingFarah TalibNo ratings yet
- Section 12.1-12.2Document18 pagesSection 12.1-12.2Ahmed AlouhNo ratings yet
- Introduction To Electrical Engineering I EE111 Introduction To Electrical Engineering I EE111Document1 pageIntroduction To Electrical Engineering I EE111 Introduction To Electrical Engineering I EE111Abdul Halil AbdullahNo ratings yet
- Linear Vs Non-Linear AnalysisDocument6 pagesLinear Vs Non-Linear AnalysisMithun DNo ratings yet
- APPLICATIONS Metal Forming Press StampingDocument3 pagesAPPLICATIONS Metal Forming Press Stamping张淼No ratings yet
- RTN 950 Commissioning Guide (U2000) - (V100R002C00 - 04) PDFDocument177 pagesRTN 950 Commissioning Guide (U2000) - (V100R002C00 - 04) PDFckiljaNo ratings yet
- SensorDocument862 pagesSensorhau nguyenNo ratings yet
- 203S.HW 11 (14.3) (Homework) : Current Score Due DateDocument13 pages203S.HW 11 (14.3) (Homework) : Current Score Due DatexvNo ratings yet
- Optimization of Rotary Drilling Configuration and Techniques - Mark BakerDocument57 pagesOptimization of Rotary Drilling Configuration and Techniques - Mark BakerSusy Serrano FlawersNo ratings yet
- Chemical FormulaeDocument4 pagesChemical FormulaeShasha FarzanaNo ratings yet
- Renold Brand Chain Tables PDFDocument4 pagesRenold Brand Chain Tables PDFAltino MangiwaNo ratings yet
- Rechargeable Sealed Lead-Acid Battery: 6 Volt 4.5 Amp. HrsDocument2 pagesRechargeable Sealed Lead-Acid Battery: 6 Volt 4.5 Amp. HrsJuan EsNo ratings yet
- Full Gravityand Inertia Printedrevised 2Document8 pagesFull Gravityand Inertia Printedrevised 2satish acu-healerNo ratings yet
- Chemical Bonding and Molecular StructureDocument14 pagesChemical Bonding and Molecular Structurepatelkrupa798No ratings yet
- Thermodynamics Week 2Document13 pagesThermodynamics Week 2Mohammad Rashik Zaman100% (1)
- Mathematical Association of AmericaDocument5 pagesMathematical Association of AmericaPatrick WuNo ratings yet
- Classifying Visual Defects in Glass-Reinforced Plastic Laminate PartsDocument24 pagesClassifying Visual Defects in Glass-Reinforced Plastic Laminate PartsDANIEL GUERRERO SANCHEZNo ratings yet
- Engineering Mechanics: Statics in SI Units, 12e: FrictionDocument31 pagesEngineering Mechanics: Statics in SI Units, 12e: FrictionZiyad AlharbiNo ratings yet
- HPLC Primaide 400 BarDocument11 pagesHPLC Primaide 400 BarMerck Millipore Brasil - Lab Supply BrasilNo ratings yet
- How To Use A Torque WrenchDocument12 pagesHow To Use A Torque WrenchVipuchit Sirikhemaporn100% (2)
- Science X - Full Test 01Document6 pagesScience X - Full Test 01Aman9692No ratings yet
- SonalDocument348 pagesSonalPallavi PandeyNo ratings yet





























































































Download as pdf or txt
You might also like
- ENEN-635 Environmental Modeling Lab-3 Report - Rev1Document10 pagesENEN-635 Environmental Modeling Lab-3 Report - Rev1Muhammad NaeemNo ratings yet
- Temperature Distribution PDFDocument5 pagesTemperature Distribution PDFEstefania ArizagaNo ratings yet
- 1 s2.0 S0032579119445367 MainDocument9 pages1 s2.0 S0032579119445367 Mainjimmy kNo ratings yet
- The Critical Importance of Incubation Temperature: Nick A. FrenchDocument5 pagesThe Critical Importance of Incubation Temperature: Nick A. FrenchThE BoNg TeChniCaLNo ratings yet
- Lin 2005Document7 pagesLin 2005Jake Floyd G. FabianNo ratings yet
- Heat and Moisture Production of Broilers Kept On Straw BeddingDocument11 pagesHeat and Moisture Production of Broilers Kept On Straw BeddingAlejandro OviedoNo ratings yet
- Body Temperatures in Free - Ying PigeonsDocument5 pagesBody Temperatures in Free - Ying PigeonsDummy DonNo ratings yet
- Chick Quality2Document12 pagesChick Quality2Dani GarnidaNo ratings yet
- Effect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualityDocument6 pagesEffect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualitySebastian RodriguezNo ratings yet
- (Apis Mellifera) Foraging in The Sonoran Desert: Temperature Regulation of Honey BeesDocument16 pages(Apis Mellifera) Foraging in The Sonoran Desert: Temperature Regulation of Honey BeeskarlaNo ratings yet
- Arch Dis Child 1975 Aynsley Green 215 9Document6 pagesArch Dis Child 1975 Aynsley Green 215 9syncmaster1234567890No ratings yet
- Thermoregulation in PoultryDocument7 pagesThermoregulation in PoultryDilip GuptaNo ratings yet
- Erjsp.2011.51.56 Vol ConsiderationDocument6 pagesErjsp.2011.51.56 Vol Considerationoluchukwu u.No ratings yet
- 1 s2.0 S0032579119439527 MainDocument11 pages1 s2.0 S0032579119439527 Mainkumpul tugasNo ratings yet
- Conditions For Cold Stress Development in Dairy Cattle Kept in Free Stall Barn During Severe FrostsDocument8 pagesConditions For Cold Stress Development in Dairy Cattle Kept in Free Stall Barn During Severe FrostsMuhammad Firdaus FebriansyahNo ratings yet
- A Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsDocument8 pagesA Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsSebastian RodriguezNo ratings yet
- Seasonal Temperature Effects and Predicting Development Rates of Marine Copepod EggsDocument7 pagesSeasonal Temperature Effects and Predicting Development Rates of Marine Copepod EggsJordan IsmaelNo ratings yet
- Wasmen HEAT STRESSDocument4 pagesWasmen HEAT STRESSByki HaqiNo ratings yet
- Modelling of Heat and Mass Transfer Processes in NeonatologyDocument12 pagesModelling of Heat and Mass Transfer Processes in NeonatologySai Santhosh ManepallyNo ratings yet
- Biocarbon Production From Hungarian Sunflower ShellsDocument5 pagesBiocarbon Production From Hungarian Sunflower ShellsMuhamad SuharNo ratings yet
- 2009.10.13 Design and Performance Evaluation of Automatic Egg IncubatorDocument13 pages2009.10.13 Design and Performance Evaluation of Automatic Egg IncubatorJohn Taulo100% (1)
- Regulacion de Temperatura AvesDocument32 pagesRegulacion de Temperatura AvesSam Lopez LopezNo ratings yet
- 08a8 PDFDocument18 pages08a8 PDFstteevennnNo ratings yet
- 1 s2.0 S0022474X14000435 Main PDFDocument6 pages1 s2.0 S0022474X14000435 Main PDFMario CalderonNo ratings yet
- Journal of Stored Products Research: R. Rusinek, R. Koby 1kaDocument6 pagesJournal of Stored Products Research: R. Rusinek, R. Koby 1kaMario CalderonNo ratings yet
- Brood Production Increases When Artificial Heating Is Provided To Colonies of Stingless BeesDocument6 pagesBrood Production Increases When Artificial Heating Is Provided To Colonies of Stingless BeesOtaviano Carneiro Da Cunha NetoNo ratings yet
- Group-2 ChE-135 Post Lab Experiment-1Document18 pagesGroup-2 ChE-135 Post Lab Experiment-1Rievelyn MartusNo ratings yet
- Heat Transfer Process TUT (After Mid Semester)Document11 pagesHeat Transfer Process TUT (After Mid Semester)vaishnavi singh100% (1)
- Susceptibility of Broiler Chickens To Hemorrhages in Muscles: The Effect of Stock and Rearing Temperature RegimenDocument8 pagesSusceptibility of Broiler Chickens To Hemorrhages in Muscles: The Effect of Stock and Rearing Temperature RegimenEbrahim AbdullaNo ratings yet
- Analisis Dan Simulasi Distribusi Suhu Udara Pada Kandang Sapi Perah Menggunakan Computational Fluid Dynamics (CFD)Document11 pagesAnalisis Dan Simulasi Distribusi Suhu Udara Pada Kandang Sapi Perah Menggunakan Computational Fluid Dynamics (CFD)SatriantyNo ratings yet
- Specific Heat of Agricultural and Food MaterialsDocument29 pagesSpecific Heat of Agricultural and Food MaterialskqNo ratings yet
- Lumped Heat Transfer AnalysisDocument29 pagesLumped Heat Transfer AnalysisprasannaNo ratings yet
- Temperature and Heat 3Document18 pagesTemperature and Heat 3Samud MorrisNo ratings yet
- Honeybee Colony Thermoregulation - Regulatory Mechanisms and Contribution of Individuals in Dependence On Age, Location and Thermal StressDocument13 pagesHoneybee Colony Thermoregulation - Regulatory Mechanisms and Contribution of Individuals in Dependence On Age, Location and Thermal Stresslosaron437No ratings yet
- Beehive - Temperature and Humidity ControlDocument7 pagesBeehive - Temperature and Humidity ControlJoao FerreiraNo ratings yet
- Thermal Properties of Butter: Peratures Between 17 and - 35°CDocument4 pagesThermal Properties of Butter: Peratures Between 17 and - 35°CpeshoaNo ratings yet
- Investigation of Effective Thermal Conductivity Kinetics of Crust and Core Regions of Potato During Deep Fat Frying JFEDocument6 pagesInvestigation of Effective Thermal Conductivity Kinetics of Crust and Core Regions of Potato During Deep Fat Frying JFELucas PanNo ratings yet
- Acclimation To Hypothermic Incubation in Developing Chicken Embryos (Gallus Domesticus)Document10 pagesAcclimation To Hypothermic Incubation in Developing Chicken Embryos (Gallus Domesticus)Sharon TarquiniNo ratings yet
- TMP 56 BDDocument4 pagesTMP 56 BDFrontiersNo ratings yet
- Calulation of Heat Sterilization Time of Fermentation MediaDocument8 pagesCalulation of Heat Sterilization Time of Fermentation Mediahansenmike698105No ratings yet
- Kustermann1982 RespirationDocument12 pagesKustermann1982 RespirationLiên Nguyễn ThùyNo ratings yet
- ETUDE - Energic Optimisation of Foraging Honeybees - Flexible Change of Strategies in Response To Environmental ChallengesDocument8 pagesETUDE - Energic Optimisation of Foraging Honeybees - Flexible Change of Strategies in Response To Environmental Challengeslosaron437No ratings yet
- ETUDE - Contribution of Honeybee Drones of Different Age To ColonialDocument17 pagesETUDE - Contribution of Honeybee Drones of Different Age To Coloniallosaron437No ratings yet
- INTRODUCTIONDocument3 pagesINTRODUCTIONLeonessa FlorNo ratings yet
- 2 IpekDocument4 pages2 IpekDani GarnidaNo ratings yet
- Untitled DocumentDocument6 pagesUntitled DocumentmandyngoaNo ratings yet
- Heat Stress in AnimalsDocument6 pagesHeat Stress in AnimalsAnke NemirovskyNo ratings yet
- Nordica RocketDocument14 pagesNordica RocketradianNo ratings yet
- Tugas Ehan Perpindahan PanasDocument44 pagesTugas Ehan Perpindahan PanasEHANNo ratings yet
- CAT 2019 Slot 1 RC 2 How Emperor Penguins Survive Antarctica's ChillDocument4 pagesCAT 2019 Slot 1 RC 2 How Emperor Penguins Survive Antarctica's ChillMana GargiNo ratings yet
- Reanimation of Rats From Body Temperatures Between 0 and 1°c by Microwave DiathermyDocument6 pagesReanimation of Rats From Body Temperatures Between 0 and 1°c by Microwave DiathermyLJNo ratings yet
- Prediction of Chilling Times of Foods in Situations Where Evaporative Cooling Is Significant-Part 1. Method DevelopmentDocument15 pagesPrediction of Chilling Times of Foods in Situations Where Evaporative Cooling Is Significant-Part 1. Method DevelopmentMaurício de OliveiraNo ratings yet
- 2022 Computers (Artigo Dissertação)Document10 pages2022 Computers (Artigo Dissertação)marcelo.bahuti2No ratings yet
- 13 Space Heating and Cooling: Properties, Production, ApplicationsDocument1 page13 Space Heating and Cooling: Properties, Production, ApplicationsPon lathaNo ratings yet
- Tazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRDocument6 pagesTazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRSebastian RodriguezNo ratings yet
- ChE 0101 Final ReportDocument29 pagesChE 0101 Final ReportPaartiban PaneerselvamNo ratings yet
- Rep-Jrf 114 2 001Document16 pagesRep-Jrf 114 2 001siyopin173No ratings yet
- The Importance of Air Velocity in IncubationDocument2 pagesThe Importance of Air Velocity in Incubationamamùra maamarNo ratings yet
- Inspires Gas Temperature Un NeonatesDocument5 pagesInspires Gas Temperature Un NeonatesGuillermo MartínezNo ratings yet
- Study of The Bread Baking Process - II. Mathematical ModellingtDocument16 pagesStudy of The Bread Baking Process - II. Mathematical Modellingtmatias andradeNo ratings yet
- Heat and Moisture Transfer between Human Body and EnvironmentFrom EverandHeat and Moisture Transfer between Human Body and EnvironmentNo ratings yet
- Robot RaplimDocument1 pageRobot RaplimSebastian Rodriguez100% (1)
- Diagrama2 PDFDocument1 pageDiagrama2 PDFSebastian RodriguezNo ratings yet
- Food Control: A A B C BDocument9 pagesFood Control: A A B C BSebastian RodriguezNo ratings yet
- Diagrama4 PDFDocument1 pageDiagrama4 PDFSebastian RodriguezNo ratings yet
- Experimental Study of The Dependence of Embryonic Development of Trachurus Trachurus Eggs On TemperatureDocument8 pagesExperimental Study of The Dependence of Embryonic Development of Trachurus Trachurus Eggs On TemperatureSebastian RodriguezNo ratings yet
- Tazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRDocument6 pagesTazawa-Nakagawa1985 Article ResponseOfEggTemperatureHeartRSebastian RodriguezNo ratings yet
- Using Multinomial Models To Analyse Data From Iberian Sardine Egg Incubation Experiments: A Comparison With Traditional TechniquesDocument9 pagesUsing Multinomial Models To Analyse Data From Iberian Sardine Egg Incubation Experiments: A Comparison With Traditional TechniquesSebastian RodriguezNo ratings yet
- Effect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualityDocument6 pagesEffect of Egg Weight and Position Relative To Incubator Fan On Broiler Hatchability and Chick QualitySebastian RodriguezNo ratings yet
- A Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsDocument8 pagesA Long-Term Study of The Environment Within A Tunnel Incubator For Turkey EggsSebastian RodriguezNo ratings yet
- Module 5 - Newtons LawsDocument11 pagesModule 5 - Newtons LawsKrizel CalasaraNo ratings yet
- 3.3a VSEPR Theory PDFDocument21 pages3.3a VSEPR Theory PDFSarvesh Kanaujia100% (8)
- Specular Reflection FTIR: A Non-Contact Method For Analyzing Coatings On Photographs and Other Cultural MaterialsDocument15 pagesSpecular Reflection FTIR: A Non-Contact Method For Analyzing Coatings On Photographs and Other Cultural MaterialsAndres Camilo Gonzalez RomeroNo ratings yet
- Good..... Analysis and Controller Design For Standalone VSIs in Synchronous Reference FrameDocument10 pagesGood..... Analysis and Controller Design For Standalone VSIs in Synchronous Reference FrameAQEEL ANWARNo ratings yet
- Arduino CertificationDocument8 pagesArduino Certificationhack reportNo ratings yet
- Practive Problems Prepared By: Engr. Dean Domenique E. ClidoroDocument11 pagesPractive Problems Prepared By: Engr. Dean Domenique E. ClidoroGrAxieQuidOrNo ratings yet
- Airgun ArraysDocument21 pagesAirgun ArraysGianni PavanNo ratings yet
- Piperazine Impregnation On Zeolite 13X As A Novel Adsorbent For CO Capture: Experimental and ModelingDocument18 pagesPiperazine Impregnation On Zeolite 13X As A Novel Adsorbent For CO Capture: Experimental and ModelingFarah TalibNo ratings yet
- Section 12.1-12.2Document18 pagesSection 12.1-12.2Ahmed AlouhNo ratings yet
- Introduction To Electrical Engineering I EE111 Introduction To Electrical Engineering I EE111Document1 pageIntroduction To Electrical Engineering I EE111 Introduction To Electrical Engineering I EE111Abdul Halil AbdullahNo ratings yet
- Linear Vs Non-Linear AnalysisDocument6 pagesLinear Vs Non-Linear AnalysisMithun DNo ratings yet
- APPLICATIONS Metal Forming Press StampingDocument3 pagesAPPLICATIONS Metal Forming Press Stamping张淼No ratings yet
- RTN 950 Commissioning Guide (U2000) - (V100R002C00 - 04) PDFDocument177 pagesRTN 950 Commissioning Guide (U2000) - (V100R002C00 - 04) PDFckiljaNo ratings yet
- SensorDocument862 pagesSensorhau nguyenNo ratings yet
- 203S.HW 11 (14.3) (Homework) : Current Score Due DateDocument13 pages203S.HW 11 (14.3) (Homework) : Current Score Due DatexvNo ratings yet
- Optimization of Rotary Drilling Configuration and Techniques - Mark BakerDocument57 pagesOptimization of Rotary Drilling Configuration and Techniques - Mark BakerSusy Serrano FlawersNo ratings yet
- Chemical FormulaeDocument4 pagesChemical FormulaeShasha FarzanaNo ratings yet
- Renold Brand Chain Tables PDFDocument4 pagesRenold Brand Chain Tables PDFAltino MangiwaNo ratings yet
- Rechargeable Sealed Lead-Acid Battery: 6 Volt 4.5 Amp. HrsDocument2 pagesRechargeable Sealed Lead-Acid Battery: 6 Volt 4.5 Amp. HrsJuan EsNo ratings yet
- Full Gravityand Inertia Printedrevised 2Document8 pagesFull Gravityand Inertia Printedrevised 2satish acu-healerNo ratings yet
- Chemical Bonding and Molecular StructureDocument14 pagesChemical Bonding and Molecular Structurepatelkrupa798No ratings yet
- Thermodynamics Week 2Document13 pagesThermodynamics Week 2Mohammad Rashik Zaman100% (1)
- Mathematical Association of AmericaDocument5 pagesMathematical Association of AmericaPatrick WuNo ratings yet
- Classifying Visual Defects in Glass-Reinforced Plastic Laminate PartsDocument24 pagesClassifying Visual Defects in Glass-Reinforced Plastic Laminate PartsDANIEL GUERRERO SANCHEZNo ratings yet
- Engineering Mechanics: Statics in SI Units, 12e: FrictionDocument31 pagesEngineering Mechanics: Statics in SI Units, 12e: FrictionZiyad AlharbiNo ratings yet
- HPLC Primaide 400 BarDocument11 pagesHPLC Primaide 400 BarMerck Millipore Brasil - Lab Supply BrasilNo ratings yet
- How To Use A Torque WrenchDocument12 pagesHow To Use A Torque WrenchVipuchit Sirikhemaporn100% (2)
- Science X - Full Test 01Document6 pagesScience X - Full Test 01Aman9692No ratings yet
- SonalDocument348 pagesSonalPallavi PandeyNo ratings yet