Download as pdf or txt
You might also like
- Lsat PT 3Document36 pagesLsat PT 3scribdfree4mo100% (5)
- Dna Mutations Worksheet Key 1617Document2 pagesDna Mutations Worksheet Key 1617Cyan AbaraNo ratings yet
- Escala FAOS IAPTDocument6 pagesEscala FAOS IAPTRui Pedro SantiagoNo ratings yet
- Reiterative Signaling and Patterning During MammalianDocument11 pagesReiterative Signaling and Patterning During MammalianJhon Mario Pinto RamirezNo ratings yet
- Epithelial-Mesenchymal Signalling Regulating Tooth MorphogenesisDocument2 pagesEpithelial-Mesenchymal Signalling Regulating Tooth MorphogenesisalifNo ratings yet
- Mitsiadis 2005Document4 pagesMitsiadis 2005MAYRA ALEJANDRA BAHAMON TRUJILLONo ratings yet
- DI and DD ClassificationDocument7 pagesDI and DD ClassificationSarmad FarooqiNo ratings yet
- p63 and IRF6: Brothers in Arms Against Cleft Palate: CommentariesDocument4 pagesp63 and IRF6: Brothers in Arms Against Cleft Palate: CommentariesNadya PurwantyNo ratings yet
- Butchova Et Al 2008 Snake DentitionDocument14 pagesButchova Et Al 2008 Snake DentitionNathalia gallegoNo ratings yet
- Morphological Analyses and A Novel de Novo DLX3 Mutation Associated With Tricho - Dento-Osseous Syndrome in A Chinese FamilyDocument8 pagesMorphological Analyses and A Novel de Novo DLX3 Mutation Associated With Tricho - Dento-Osseous Syndrome in A Chinese FamilyMia De DiosNo ratings yet
- Tooth Root FormationDocument10 pagesTooth Root FormationAthenaeum Scientific PublishersNo ratings yet
- Archives of Oral Biology: Original ArticleDocument4 pagesArchives of Oral Biology: Original ArticleKaritoCobosNo ratings yet
- Nguyen 2013Document5 pagesNguyen 2013Biologija BiologyNo ratings yet
- MoleculaDocument114 pagesMoleculaDilla Novia AmrilaniNo ratings yet
- Ectodysplasin A (EDA)Document9 pagesEctodysplasin A (EDA)Alex CasarrubiasNo ratings yet
- Paper Biofisica 1Document7 pagesPaper Biofisica 1Mauro MarchettoNo ratings yet
- The Role of Nonhomologous End Joining CoDocument11 pagesThe Role of Nonhomologous End Joining Cokilli999No ratings yet
- Artigo 1Document7 pagesArtigo 1IsabelNo ratings yet
- ZhangDocument11 pagesZhangAJMRNo ratings yet
- Epigenetic Regulation of The Immune SystemDocument8 pagesEpigenetic Regulation of The Immune SystemDidacNo ratings yet
- Huang2012 PDFDocument13 pagesHuang2012 PDFtsth4ckNo ratings yet
- 1 s2.0 000399699090093P MainDocument9 pages1 s2.0 000399699090093P MainNancyLiaoNo ratings yet
- Ohi (2006)Document6 pagesOhi (2006)Bakhshi Dlshad MajeedNo ratings yet
- Cobourne (1999) The Genetic Control of Early OdontogenesisDocument0 pagesCobourne (1999) The Genetic Control of Early OdontogenesisMaja Maja BułkaNo ratings yet
- Dento-Craniofacial Phenotypes and Underlying Molecular Mechanisms in Hypohidrotic Ectodermal Dysplasia (Hed) : A ReviewDocument11 pagesDento-Craniofacial Phenotypes and Underlying Molecular Mechanisms in Hypohidrotic Ectodermal Dysplasia (Hed) : A ReviewYusuf DiansyahNo ratings yet
- The Genetic Basis of Normal and Abnormal Craniofacial Development. Acta Odontol Scand, 56 321-325, 1998Document6 pagesThe Genetic Basis of Normal and Abnormal Craniofacial Development. Acta Odontol Scand, 56 321-325, 1998Muhammad AsimNo ratings yet
- Zebrafish As A Model To Study The Role of DNA Methylation in Environmental ToxicologyDocument15 pagesZebrafish As A Model To Study The Role of DNA Methylation in Environmental ToxicologyTiago TorresNo ratings yet
- LeuO Regula Negativamente SPI1Document13 pagesLeuO Regula Negativamente SPI1DiegoNo ratings yet
- Multiple Bilateral Taurodontism. A Case ReportDocument3 pagesMultiple Bilateral Taurodontism. A Case ReportJing XueNo ratings yet
- Bras PereiraMOD08Document13 pagesBras PereiraMOD08Fernando CasaresNo ratings yet
- The Genetic Basis of Dental Anomalies and Its Relation To OrthodonticsDocument5 pagesThe Genetic Basis of Dental Anomalies and Its Relation To Orthodonticsyuniarita eka putriNo ratings yet
- Hum. Mol. Genet. 2001 Rajpar 1673 7Document6 pagesHum. Mol. Genet. 2001 Rajpar 1673 7Nadya PurwantyNo ratings yet
- Msx1/Bmp4 Genetic Pathway Regulates Mammalian Alveolar Bone Formation Via Induction of Dlx5 and Cbfa1Document11 pagesMsx1/Bmp4 Genetic Pathway Regulates Mammalian Alveolar Bone Formation Via Induction of Dlx5 and Cbfa1Laura Putri TraviraNo ratings yet
- Genetics of AbnormalitiesDocument11 pagesGenetics of AbnormalitiesLjubomirErdoglijaNo ratings yet
- Articulo 5Document8 pagesArticulo 5FernierNo ratings yet
- Epigenetics - A New Frontier in DentistryDocument11 pagesEpigenetics - A New Frontier in DentistryDiego Andres Hincapie HerreraNo ratings yet
- 2018 Article 342Document9 pages2018 Article 342vivigaitanNo ratings yet
- Accepted Manuscript: 10.1016/j.cortex.2015.07.004Document20 pagesAccepted Manuscript: 10.1016/j.cortex.2015.07.004pokharelriwaj82No ratings yet
- Structural and Functional ChangesDocument9 pagesStructural and Functional ChangesMAGALI GONZÁLEZ RODRÍGUEZNo ratings yet
- MRI Analysis of An Inherited Speech and Language DDocument15 pagesMRI Analysis of An Inherited Speech and Language Dcarmooh14No ratings yet
- Thesleff 2018Document5 pagesThesleff 2018jalozanorNo ratings yet
- Lopes Casares DB10Document12 pagesLopes Casares DB10Fernando CasaresNo ratings yet
- Transkripsiyon FaktörleriDocument16 pagesTranskripsiyon FaktörleriEhed AymazNo ratings yet
- Vip-2010.cytidine Deaminases AIDing DNA DemethylationDocument9 pagesVip-2010.cytidine Deaminases AIDing DNA Demethylationhualin2010No ratings yet
- Dental Manifestations of Dermatologic Conditions: Journal of The American Academy of Dermatology December 2008Document11 pagesDental Manifestations of Dermatologic Conditions: Journal of The American Academy of Dermatology December 2008faty basalamahNo ratings yet
- Pi Is 0092867418301065Document19 pagesPi Is 0092867418301065Khushi TalaviyaNo ratings yet
- DGI TIPE 2 BDocument4 pagesDGI TIPE 2 BYurika HandayaniNo ratings yet
- Novel Human MutationDocument8 pagesNovel Human Mutationnana syahlaNo ratings yet
- Developmental Biology: Abigail M. Anderson, Brandon P. Weasner, Bonnie M. Weasner, Justin P. KumarDocument11 pagesDevelopmental Biology: Abigail M. Anderson, Brandon P. Weasner, Bonnie M. Weasner, Justin P. KumartinaNo ratings yet
- Paper 5Document9 pagesPaper 5fjidjitopNo ratings yet
- Practical Report About Mutant Type of Drosophila Melanogaster - Yolanda AnggraeniDocument7 pagesPractical Report About Mutant Type of Drosophila Melanogaster - Yolanda Anggraeniyolanda anggraeniNo ratings yet
- Molecular and Cellular Mechanisms of Tooth Development...Document15 pagesMolecular and Cellular Mechanisms of Tooth Development...dophannhuystgNo ratings yet
- Liudmyla Tsyba, Oleksii Nikolaienko, Oleksandr Dergai, Mykola Dergai, Olga Novokhatska, Inessa Skrypkina, Alla RynditchDocument9 pagesLiudmyla Tsyba, Oleksii Nikolaienko, Oleksandr Dergai, Mykola Dergai, Olga Novokhatska, Inessa Skrypkina, Alla RynditchSergeat18BNo ratings yet
- LivakDocument24 pagesLivakibrahima1968No ratings yet
- Genetics in OrthodonticsDocument56 pagesGenetics in OrthodonticsSadhu AbhijeetNo ratings yet
- Expression and Localization of Laminin-5 Subunits in The Mouse IncisorDocument7 pagesExpression and Localization of Laminin-5 Subunits in The Mouse Incisornovi putri ayuNo ratings yet
- DNA Meth in Social Spiders, Liu Et Al, 2019Document17 pagesDNA Meth in Social Spiders, Liu Et Al, 2019kyudionNo ratings yet
- Contacto Lengua A Paldar Durante El Habla en Sujetos Con y Sin Interposicon Lingual 2006Document5 pagesContacto Lengua A Paldar Durante El Habla en Sujetos Con y Sin Interposicon Lingual 2006Romi Sepúlveda SaldiviaNo ratings yet
- Grand Challenges in Comparative Tooth Biology: SymposiumDocument18 pagesGrand Challenges in Comparative Tooth Biology: SymposiumSamuelHpNo ratings yet
- Can Waardenburg Syndrome Type 2 Be Explained by Epigenetic Mosaicism?Document3 pagesCan Waardenburg Syndrome Type 2 Be Explained by Epigenetic Mosaicism?Iso MogoNo ratings yet
- 2010 - UzDocument8 pages2010 - UzCarlNo ratings yet
- YangDocument13 pagesYangAJMRNo ratings yet
- Herrera 2014Document15 pagesHerrera 2014Jhon Mario Pinto RamirezNo ratings yet
- Williams 2007Document19 pagesWilliams 2007Jhon Mario Pinto RamirezNo ratings yet
- Foot Notes The Social Implications of Polydactyly and Foot-RelatedDocument24 pagesFoot Notes The Social Implications of Polydactyly and Foot-RelatedJhon Mario Pinto RamirezNo ratings yet
- Plagiarism Scan Report: Content Checked For PlagiarismDocument1 pagePlagiarism Scan Report: Content Checked For PlagiarismJhon Mario Pinto RamirezNo ratings yet
- GetpdfsplitzipdocumentDocument39 pagesGetpdfsplitzipdocumentJhon Mario Pinto RamirezNo ratings yet
- HIstoria de Los Chami Zuluaga 1988Document133 pagesHIstoria de Los Chami Zuluaga 1988Jhon Mario Pinto RamirezNo ratings yet
- Reiterative Signaling and Patterning During MammalianDocument11 pagesReiterative Signaling and Patterning During MammalianJhon Mario Pinto RamirezNo ratings yet
- Hovorakova 2007Document8 pagesHovorakova 2007Jhon Mario Pinto RamirezNo ratings yet
- Genetic Basis of Tooth Development and Dental DefectsDocument5 pagesGenetic Basis of Tooth Development and Dental DefectsJhon Mario Pinto RamirezNo ratings yet
- Evolutionary Modification of Development in Mammals TeethDocument5 pagesEvolutionary Modification of Development in Mammals TeethJhon Mario Pinto RamirezNo ratings yet
- A Gene Network Model Accounting ForDocument5 pagesA Gene Network Model Accounting ForJhon Mario Pinto RamirezNo ratings yet
- GFP TaggingDocument6 pagesGFP TaggingwaterdolNo ratings yet
- MCQ Breeding and Farm AnimalsDocument10 pagesMCQ Breeding and Farm AnimalsvishnuNo ratings yet
- Review of Literature of Medicinal PlantsDocument8 pagesReview of Literature of Medicinal Plantsgrgdazukg100% (1)
- Covid PGXDocument13 pagesCovid PGXDyahsekarayupudak wangiNo ratings yet
- Hungitons DiseaseDocument267 pagesHungitons DiseasePeter Osundwa Kiteki100% (1)
- Word Parts Dictionary Prefixes Suffixes Roots and Combining Forms.1 106Document106 pagesWord Parts Dictionary Prefixes Suffixes Roots and Combining Forms.1 106Ame QP100% (1)
- Bruce Lipton InterviewDocument28 pagesBruce Lipton InterviewDr Ron100% (2)
- Biomedicines 08 00301 v2Document24 pagesBiomedicines 08 00301 v2d.ramadhanNo ratings yet
- PB Using A Spirometer To Investigate Human Lung Function SsDocument6 pagesPB Using A Spirometer To Investigate Human Lung Function SsMohammad Shahidullah ChowdhuryNo ratings yet
- GMO Myths and TruthsDocument123 pagesGMO Myths and TruthsChildren Of Vietnam Veterans Health Alliance100% (2)
- MutationsDocument29 pagesMutationsiariajay100% (1)
- Introduction To Biological PsychologyDocument23 pagesIntroduction To Biological PsychologyBhagyashree radeNo ratings yet
- Tuberculinum PDFDocument4 pagesTuberculinum PDFadi swara100% (1)
- Cutaneous Manifestations of GIT and Liver DiseasesDocument48 pagesCutaneous Manifestations of GIT and Liver DiseasesSippy Iqbal MemonNo ratings yet
- Art 5Document28 pagesArt 5sheheryar.sixsigmatravelsNo ratings yet
- Zoology 100 Notes 1Document8 pagesZoology 100 Notes 1Bethany Jane Ravelo IsidroNo ratings yet
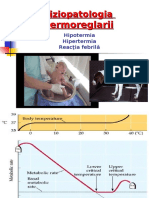
- Documents - MX Fiziopatologia Termoreglarii Fiziopatologia Termoreglarii Hipotermia Hipertermia Reactia Febrila 10-12-2006Document42 pagesDocuments - MX Fiziopatologia Termoreglarii Fiziopatologia Termoreglarii Hipotermia Hipertermia Reactia Febrila 10-12-2006AndreiNo ratings yet
- Buku Stem Cells From Basic Research To TherapyDocument670 pagesBuku Stem Cells From Basic Research To Therapyaudiary pramesti100% (1)
- NRDPUlcerative Colitis PDFDocument21 pagesNRDPUlcerative Colitis PDFLuminita HutanuNo ratings yet
- List of Eponyms in MedicineDocument12 pagesList of Eponyms in MedicinePavan V RaiNo ratings yet
- Cloning Tech Guide PDFDocument40 pagesCloning Tech Guide PDFLuis Fernando Soto UgaldiNo ratings yet
- AIIMS 2019 Biology Sample Question PaperDocument10 pagesAIIMS 2019 Biology Sample Question PapermisostudyNo ratings yet
- Extending Mendelian Genetics For Two or More GenesDocument27 pagesExtending Mendelian Genetics For Two or More GenesGerald GuevarraNo ratings yet
- Abdel-Moneim (2007) PDFDocument24 pagesAbdel-Moneim (2007) PDFinsaniachusnaNo ratings yet
- Molecules, of The Immune SystemDocument39 pagesMolecules, of The Immune SystemFaith MbasaniNo ratings yet
- A Nonsynonymous Single Nucleotide Variant of The Tumor-Related Gene ADAM15 in Colon Cancer and Its Impact On Cell Adhesion PhenotypeDocument9 pagesA Nonsynonymous Single Nucleotide Variant of The Tumor-Related Gene ADAM15 in Colon Cancer and Its Impact On Cell Adhesion Phenotypeaya younesNo ratings yet
- Case Report Pediatric Von&JepDocument23 pagesCase Report Pediatric Von&JepFarizan NurmushoffaNo ratings yet