
17 PVAT (Gao)
17 PVAT (Gao)
You might also like
- KeNotes 2017 PDFDocument106 pagesKeNotes 2017 PDFDENTAL REVIEWER ONLY100% (3)
- Gel Polish and Dry Manicure Manual EDITABLEDocument56 pagesGel Polish and Dry Manicure Manual EDITABLECristina Gradinari100% (1)
- CVS Embryology Questions and Study Guide - Quizlet Flashcards by Hugo - OxfordDocument5 pagesCVS Embryology Questions and Study Guide - Quizlet Flashcards by Hugo - OxfordAzizNo ratings yet
- Jvms 83 997Document7 pagesJvms 83 997Blanca A SerranoNo ratings yet
- Birukova MicrovascRes 2013Document8 pagesBirukova MicrovascRes 2013useretailNo ratings yet
- Central Role For Aldose Reductase Pathway in Myocardial Ischemic InjuryDocument8 pagesCentral Role For Aldose Reductase Pathway in Myocardial Ischemic Injurysinta mahastutiNo ratings yet
- 416 FullDocument9 pages416 FullFidia Hanan asystNo ratings yet
- A20Comparison20between20the20Effects20of20Hydrophobic20 PDFDocument12 pagesA20Comparison20between20the20Effects20of20Hydrophobic20 PDFnasir uddinNo ratings yet
- European Journal of PharmacologyDocument9 pagesEuropean Journal of PharmacologyMarcelo Pires de OliveiraNo ratings yet
- NIH Public Access: Author ManuscriptDocument19 pagesNIH Public Access: Author Manuscriptaldo123456789No ratings yet
- Toxicology and Applied Pharmacology: Chieko Saito, John J. Lemasters, Hartmut JaeschkeDocument10 pagesToxicology and Applied Pharmacology: Chieko Saito, John J. Lemasters, Hartmut JaeschkezaifNo ratings yet
- MJW 011Document8 pagesMJW 011eeeeNo ratings yet
- Cardioproteccion Frente Isquemia 2008Document8 pagesCardioproteccion Frente Isquemia 2008cumbredinNo ratings yet
- Journal of Functional Foods: Qinlu Lin, Wang Liao, Jie Bai, Wei Wu, Jianping WuDocument6 pagesJournal of Functional Foods: Qinlu Lin, Wang Liao, Jie Bai, Wei Wu, Jianping Wusuci apsari pebriantiNo ratings yet
- Splenic Hemodynamics and Decreased Endothelial Nitric Oxide Synthase in The Spleen of Rats With Liver CirrhosisDocument9 pagesSplenic Hemodynamics and Decreased Endothelial Nitric Oxide Synthase in The Spleen of Rats With Liver CirrhosisdenisegmeloNo ratings yet
- Effect of Hypoxia On Vasodilator Responses To SNAP and Levcromakalim in Guinea Pig Basilar ArteryDocument6 pagesEffect of Hypoxia On Vasodilator Responses To SNAP and Levcromakalim in Guinea Pig Basilar ArteryAnon0123No ratings yet
- NADH Supplementation Decreases Pinacidil-Primed IK (ATP) in Ventricular Cardiomyocytes by Increasing Intracellular ATPDocument6 pagesNADH Supplementation Decreases Pinacidil-Primed IK (ATP) in Ventricular Cardiomyocytes by Increasing Intracellular ATPmehmetozsavciNo ratings yet
- Jurnal Internasional - Vas DeferensDocument11 pagesJurnal Internasional - Vas DeferensAnggita MelaniNo ratings yet
- Stress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityDocument18 pagesStress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityCony GSNo ratings yet
- Fauconnier2007 Insulin TRPC3Document10 pagesFauconnier2007 Insulin TRPC3Giulia AndreeaNo ratings yet
- Joung2007-BM Mononuclear Cells - MIDocument11 pagesJoung2007-BM Mononuclear Cells - MIiulia andreeaNo ratings yet
- Art 1 en Dot HeliumDocument11 pagesArt 1 en Dot HeliumMiss_PiggyNo ratings yet
- KCL Evokes Contraction of Airway Smooth Muscle Via Activation of Rhoa and Rho-KinaseDocument7 pagesKCL Evokes Contraction of Airway Smooth Muscle Via Activation of Rhoa and Rho-KinaseElias CampuzanoNo ratings yet
- Lamotrigina y VejezDocument6 pagesLamotrigina y VejezJosé Luis SHNo ratings yet
- Pastor 1997Document8 pagesPastor 1997Alexsandro Nicolay PontoanNo ratings yet
- Exercise-Induced Rhabdomyolysis Mechanisms and PreventionDocument11 pagesExercise-Induced Rhabdomyolysis Mechanisms and PreventionMarlene PgNo ratings yet
- The cardioprotective effect of astaxanthin against isoprenaline-induced myocardial injury in rats: involvement of TLR4/NF-κB signaling pathwayDocument7 pagesThe cardioprotective effect of astaxanthin against isoprenaline-induced myocardial injury in rats: involvement of TLR4/NF-κB signaling pathwayMennatallah AliNo ratings yet
- Antihypertensive and Neuroprotective Effects of Astaxanthin in Experimental AnimalsDocument6 pagesAntihypertensive and Neuroprotective Effects of Astaxanthin in Experimental Animalsarif fikriNo ratings yet
- The Chemistry of Thiosulfate and Vascular Calcification: W. Charles O'Neill and Kenneth I. HardcastleDocument6 pagesThe Chemistry of Thiosulfate and Vascular Calcification: W. Charles O'Neill and Kenneth I. HardcastleZakki LabiebNo ratings yet
- Seminario 5 Degradacion de Proteinas 2012Document9 pagesSeminario 5 Degradacion de Proteinas 2012pau_sebNo ratings yet
- TMP 75 AADocument9 pagesTMP 75 AAFrontiersNo ratings yet
- Takeda 2007 Spironolakton Pada MI TikusDocument11 pagesTakeda 2007 Spironolakton Pada MI TikusauliaNo ratings yet
- TMP C32Document11 pagesTMP C32FrontiersNo ratings yet
- Marchant2022 CardiacPacemakerAPDocument7 pagesMarchant2022 CardiacPacemakerAPsabrinasameja75No ratings yet
- Perivascular Adipose Tissue 2023 CDocument7 pagesPerivascular Adipose Tissue 2023 Cmiguel contrerasNo ratings yet
- 1 s2.0 S0014299903027250 MainDocument8 pages1 s2.0 S0014299903027250 MainZulfiani SyachbaniahNo ratings yet
- 2020 Sanjeev Physiology International 2020Document15 pages2020 Sanjeev Physiology International 2020maloymandalNo ratings yet
- Docosahexaenoic Acid Inhibits Protein Kinase C Translocation/activation and Cardiac Hypertrophy in Rat CardiomyocytesDocument8 pagesDocosahexaenoic Acid Inhibits Protein Kinase C Translocation/activation and Cardiac Hypertrophy in Rat CardiomyocytesStepss StepsNo ratings yet
- Resistencia A InfartosDocument6 pagesResistencia A InfartosJosé Luis SHNo ratings yet
- P2X R-Mediated Ca - Independent - Serine Release Via Pannexin-1 of The P2X R-Pannexin-1 Complex in AstrocytesDocument17 pagesP2X R-Mediated Ca - Independent - Serine Release Via Pannexin-1 of The P2X R-Pannexin-1 Complex in AstrocytesjavillusNo ratings yet
- Statin-Induced Myopathy in Skeletal Muscle: The Role of ExerciseDocument9 pagesStatin-Induced Myopathy in Skeletal Muscle: The Role of Exercisesita laodikaNo ratings yet
- Effect of Phosphocreatine and EthylmethyDocument2 pagesEffect of Phosphocreatine and Ethylmethymercuryboss90No ratings yet
- Cardioprotector 2004Document10 pagesCardioprotector 2004cumbredinNo ratings yet
- Research Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsDocument12 pagesResearch Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsCarlos LabradaNo ratings yet
- Vol-1, Issue 7 Paper (5) Page 26-31Document7 pagesVol-1, Issue 7 Paper (5) Page 26-31IJMSHCNo ratings yet
- Acido Ursolico NNDocument5 pagesAcido Ursolico NNDIANA PATRICIA MOLANO PEREZNo ratings yet
- Volatile Anaesthetic Myocardial Protection: A Review of The Current LiteratureDocument5 pagesVolatile Anaesthetic Myocardial Protection: A Review of The Current Literatureserena7205No ratings yet
- SSRN Id4401700Document94 pagesSSRN Id4401700Milan Petrik100% (1)
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDocument8 pagesAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshNo ratings yet
- Nai Et Al 2017 Protective Effect of Astaxanthin On Acute Cerebral Infarction in RatsDocument8 pagesNai Et Al 2017 Protective Effect of Astaxanthin On Acute Cerebral Infarction in RatsMennatallah AliNo ratings yet
- Efectos Del Ultrasonido en FibroblastosDocument7 pagesEfectos Del Ultrasonido en FibroblastosErika VillacisNo ratings yet
- Vasopressin in Septic ShockDocument8 pagesVasopressin in Septic ShockVaishnavi SheteNo ratings yet
- 1 s2.0 S0925443923001850 MainDocument11 pages1 s2.0 S0925443923001850 Mainviventius yoshuaNo ratings yet
- Quercetin HypertensiveDocument8 pagesQuercetin HypertensiveAprilihardini LaksmiNo ratings yet
- The Role of The NaDocument13 pagesThe Role of The NamrtiborhornyikNo ratings yet
- Toksik Liver 9Document13 pagesToksik Liver 9Mita SilviyaniNo ratings yet
- Georgescu2013 PDFDocument9 pagesGeorgescu2013 PDFILHAM 'K FOR ALLNo ratings yet
- Expresi PeriostinDocument5 pagesExpresi Periostinkhoirul imaNo ratings yet
- Wendy W. Wu, C. Savio Chan, D. James Surmeier and John F. DisterhoftDocument13 pagesWendy W. Wu, C. Savio Chan, D. James Surmeier and John F. DisterhoftFedrmNo ratings yet
- Effect of Rosiglitazone On RabbiDocument7 pagesEffect of Rosiglitazone On Rabbinasirasna899No ratings yet
- 2007-Effects and Mechanisms of Electroacupuncture at PC6 On Frequency of Transient Lower Esophageal Sphincter Relaxation in Cats PDFDocument8 pages2007-Effects and Mechanisms of Electroacupuncture at PC6 On Frequency of Transient Lower Esophageal Sphincter Relaxation in Cats PDFvictoregvNo ratings yet
- Involvement of Parasympathetic Pelvic Efferent Pathway in Psychological Stress-Induced DefecationDocument10 pagesInvolvement of Parasympathetic Pelvic Efferent Pathway in Psychological Stress-Induced DefecationHrvoje JakovacNo ratings yet
- Multi-scale Extracellular Matrix Mechanics and MechanobiologyFrom EverandMulti-scale Extracellular Matrix Mechanics and MechanobiologyYanhang ZhangNo ratings yet
- Embryology of The Heart and The Great VesselsDocument66 pagesEmbryology of The Heart and The Great VesselsNavya HegdeNo ratings yet
- Overhead Transparency Human AtlasDocument34 pagesOverhead Transparency Human Atlasdr tamerNo ratings yet
- Science - G9 - Week 2 (Lessons-4-6)Document46 pagesScience - G9 - Week 2 (Lessons-4-6)Rhyan Zero-four BaluyutNo ratings yet
- Through The Decades (Beta) - Blocker Use and Outcomes in Acute Coronary Syndromes.Document8 pagesThrough The Decades (Beta) - Blocker Use and Outcomes in Acute Coronary Syndromes.Tony Miguel Saba SabaNo ratings yet
- Epidemiology, Risk Factors, Pathogenesis, and Natural History of Abdominal Aortic Aneurysm - UpToDateDocument15 pagesEpidemiology, Risk Factors, Pathogenesis, and Natural History of Abdominal Aortic Aneurysm - UpToDateALVARO ARIASNo ratings yet
- Yarber Autopsy ReportDocument7 pagesYarber Autopsy ReportSarah BatchaNo ratings yet
- GEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Document202 pagesGEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Shantanu MandalNo ratings yet
- Falenissa - Pemicu 3 Blok Etika Dan Hukum KedokteranDocument108 pagesFalenissa - Pemicu 3 Blok Etika Dan Hukum KedokteranIcarus WingsNo ratings yet
- Logiq e Datasheet PDFDocument12 pagesLogiq e Datasheet PDFrca ieftinNo ratings yet
- Doppler Echocardiography For The Small Animal Practitioner June A Boon Full ChapterDocument67 pagesDoppler Echocardiography For The Small Animal Practitioner June A Boon Full Chapterelizabeth.myers417100% (12)
- 110 TOP SURGERY Multiple Choice Questions and Answers PDF - Medical Multiple Choice Questions PDFDocument11 pages110 TOP SURGERY Multiple Choice Questions and Answers PDF - Medical Multiple Choice Questions PDFaziz0% (1)
- BrahmaiahDocument395 pagesBrahmaiahBrahmaiah UpputuriNo ratings yet
- Anatomy of The AortaDocument19 pagesAnatomy of The AortaUma Sanggari ThirunavukarasuNo ratings yet
- Measurements in Radiology Made EasyDocument213 pagesMeasurements in Radiology Made EasySllavko K. KallfaNo ratings yet
- Full Download Test Bank For Human Embryology and Developmental Biology 6th Edition Carlson PDF Full ChapterDocument36 pagesFull Download Test Bank For Human Embryology and Developmental Biology 6th Edition Carlson PDF Full Chapterjuristvesicantg7egk100% (22)
- Histologi Organ Dan Sistem Sirkulasi PoltekkesDocument28 pagesHistologi Organ Dan Sistem Sirkulasi PoltekkesJeon WonwooNo ratings yet
- State Group CCMP MCQDocument28 pagesState Group CCMP MCQGirish WaruNo ratings yet
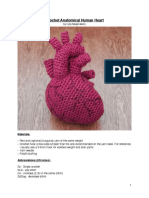
- Crochet Anatomical Human Heart: by Kyla RaaymakersDocument7 pagesCrochet Anatomical Human Heart: by Kyla RaaymakersJavier BejaranoNo ratings yet
- Anatomy Quest.Document11 pagesAnatomy Quest.Ade AlcarazNo ratings yet
- Anatomy Physiology of The BrainDocument107 pagesAnatomy Physiology of The BrainRadha RamineniNo ratings yet
- Acute Coronary Syndrome Case-Report: A U F Angeles City College of NursingDocument77 pagesAcute Coronary Syndrome Case-Report: A U F Angeles City College of NursingCarlo Joseph Intal LlacerNo ratings yet
- ITQR14Document6 pagesITQR14Vasu NallaluthanNo ratings yet
- Diaphragm AnatomyDocument19 pagesDiaphragm AnatomyasujithdrNo ratings yet
- Biology Mnemonics (Neet - Aiims - Jipmer)Document67 pagesBiology Mnemonics (Neet - Aiims - Jipmer)MyCollectionMtgNo ratings yet
- PDFDocument97 pagesPDFUmeshNo ratings yet
- Coronary Artery Disease - Case StudyDocument19 pagesCoronary Artery Disease - Case StudyJulieteeySarania100% (4)
- For Questions Number 1-5, Refer To Scenario BelowDocument48 pagesFor Questions Number 1-5, Refer To Scenario BelowPuravin Crishan VeerasamyNo ratings yet

























































































Download as pdf or txt
You might also like
- KeNotes 2017 PDFDocument106 pagesKeNotes 2017 PDFDENTAL REVIEWER ONLY100% (3)
- Gel Polish and Dry Manicure Manual EDITABLEDocument56 pagesGel Polish and Dry Manicure Manual EDITABLECristina Gradinari100% (1)
- CVS Embryology Questions and Study Guide - Quizlet Flashcards by Hugo - OxfordDocument5 pagesCVS Embryology Questions and Study Guide - Quizlet Flashcards by Hugo - OxfordAzizNo ratings yet
- Jvms 83 997Document7 pagesJvms 83 997Blanca A SerranoNo ratings yet
- Birukova MicrovascRes 2013Document8 pagesBirukova MicrovascRes 2013useretailNo ratings yet
- Central Role For Aldose Reductase Pathway in Myocardial Ischemic InjuryDocument8 pagesCentral Role For Aldose Reductase Pathway in Myocardial Ischemic Injurysinta mahastutiNo ratings yet
- 416 FullDocument9 pages416 FullFidia Hanan asystNo ratings yet
- A20Comparison20between20the20Effects20of20Hydrophobic20 PDFDocument12 pagesA20Comparison20between20the20Effects20of20Hydrophobic20 PDFnasir uddinNo ratings yet
- European Journal of PharmacologyDocument9 pagesEuropean Journal of PharmacologyMarcelo Pires de OliveiraNo ratings yet
- NIH Public Access: Author ManuscriptDocument19 pagesNIH Public Access: Author Manuscriptaldo123456789No ratings yet
- Toxicology and Applied Pharmacology: Chieko Saito, John J. Lemasters, Hartmut JaeschkeDocument10 pagesToxicology and Applied Pharmacology: Chieko Saito, John J. Lemasters, Hartmut JaeschkezaifNo ratings yet
- MJW 011Document8 pagesMJW 011eeeeNo ratings yet
- Cardioproteccion Frente Isquemia 2008Document8 pagesCardioproteccion Frente Isquemia 2008cumbredinNo ratings yet
- Journal of Functional Foods: Qinlu Lin, Wang Liao, Jie Bai, Wei Wu, Jianping WuDocument6 pagesJournal of Functional Foods: Qinlu Lin, Wang Liao, Jie Bai, Wei Wu, Jianping Wusuci apsari pebriantiNo ratings yet
- Splenic Hemodynamics and Decreased Endothelial Nitric Oxide Synthase in The Spleen of Rats With Liver CirrhosisDocument9 pagesSplenic Hemodynamics and Decreased Endothelial Nitric Oxide Synthase in The Spleen of Rats With Liver CirrhosisdenisegmeloNo ratings yet
- Effect of Hypoxia On Vasodilator Responses To SNAP and Levcromakalim in Guinea Pig Basilar ArteryDocument6 pagesEffect of Hypoxia On Vasodilator Responses To SNAP and Levcromakalim in Guinea Pig Basilar ArteryAnon0123No ratings yet
- NADH Supplementation Decreases Pinacidil-Primed IK (ATP) in Ventricular Cardiomyocytes by Increasing Intracellular ATPDocument6 pagesNADH Supplementation Decreases Pinacidil-Primed IK (ATP) in Ventricular Cardiomyocytes by Increasing Intracellular ATPmehmetozsavciNo ratings yet
- Jurnal Internasional - Vas DeferensDocument11 pagesJurnal Internasional - Vas DeferensAnggita MelaniNo ratings yet
- Stress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityDocument18 pagesStress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityCony GSNo ratings yet
- Fauconnier2007 Insulin TRPC3Document10 pagesFauconnier2007 Insulin TRPC3Giulia AndreeaNo ratings yet
- Joung2007-BM Mononuclear Cells - MIDocument11 pagesJoung2007-BM Mononuclear Cells - MIiulia andreeaNo ratings yet
- Art 1 en Dot HeliumDocument11 pagesArt 1 en Dot HeliumMiss_PiggyNo ratings yet
- KCL Evokes Contraction of Airway Smooth Muscle Via Activation of Rhoa and Rho-KinaseDocument7 pagesKCL Evokes Contraction of Airway Smooth Muscle Via Activation of Rhoa and Rho-KinaseElias CampuzanoNo ratings yet
- Lamotrigina y VejezDocument6 pagesLamotrigina y VejezJosé Luis SHNo ratings yet
- Pastor 1997Document8 pagesPastor 1997Alexsandro Nicolay PontoanNo ratings yet
- Exercise-Induced Rhabdomyolysis Mechanisms and PreventionDocument11 pagesExercise-Induced Rhabdomyolysis Mechanisms and PreventionMarlene PgNo ratings yet
- The cardioprotective effect of astaxanthin against isoprenaline-induced myocardial injury in rats: involvement of TLR4/NF-κB signaling pathwayDocument7 pagesThe cardioprotective effect of astaxanthin against isoprenaline-induced myocardial injury in rats: involvement of TLR4/NF-κB signaling pathwayMennatallah AliNo ratings yet
- Antihypertensive and Neuroprotective Effects of Astaxanthin in Experimental AnimalsDocument6 pagesAntihypertensive and Neuroprotective Effects of Astaxanthin in Experimental Animalsarif fikriNo ratings yet
- The Chemistry of Thiosulfate and Vascular Calcification: W. Charles O'Neill and Kenneth I. HardcastleDocument6 pagesThe Chemistry of Thiosulfate and Vascular Calcification: W. Charles O'Neill and Kenneth I. HardcastleZakki LabiebNo ratings yet
- Seminario 5 Degradacion de Proteinas 2012Document9 pagesSeminario 5 Degradacion de Proteinas 2012pau_sebNo ratings yet
- TMP 75 AADocument9 pagesTMP 75 AAFrontiersNo ratings yet
- Takeda 2007 Spironolakton Pada MI TikusDocument11 pagesTakeda 2007 Spironolakton Pada MI TikusauliaNo ratings yet
- TMP C32Document11 pagesTMP C32FrontiersNo ratings yet
- Marchant2022 CardiacPacemakerAPDocument7 pagesMarchant2022 CardiacPacemakerAPsabrinasameja75No ratings yet
- Perivascular Adipose Tissue 2023 CDocument7 pagesPerivascular Adipose Tissue 2023 Cmiguel contrerasNo ratings yet
- 1 s2.0 S0014299903027250 MainDocument8 pages1 s2.0 S0014299903027250 MainZulfiani SyachbaniahNo ratings yet
- 2020 Sanjeev Physiology International 2020Document15 pages2020 Sanjeev Physiology International 2020maloymandalNo ratings yet
- Docosahexaenoic Acid Inhibits Protein Kinase C Translocation/activation and Cardiac Hypertrophy in Rat CardiomyocytesDocument8 pagesDocosahexaenoic Acid Inhibits Protein Kinase C Translocation/activation and Cardiac Hypertrophy in Rat CardiomyocytesStepss StepsNo ratings yet
- Resistencia A InfartosDocument6 pagesResistencia A InfartosJosé Luis SHNo ratings yet
- P2X R-Mediated Ca - Independent - Serine Release Via Pannexin-1 of The P2X R-Pannexin-1 Complex in AstrocytesDocument17 pagesP2X R-Mediated Ca - Independent - Serine Release Via Pannexin-1 of The P2X R-Pannexin-1 Complex in AstrocytesjavillusNo ratings yet
- Statin-Induced Myopathy in Skeletal Muscle: The Role of ExerciseDocument9 pagesStatin-Induced Myopathy in Skeletal Muscle: The Role of Exercisesita laodikaNo ratings yet
- Effect of Phosphocreatine and EthylmethyDocument2 pagesEffect of Phosphocreatine and Ethylmethymercuryboss90No ratings yet
- Cardioprotector 2004Document10 pagesCardioprotector 2004cumbredinNo ratings yet
- Research Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsDocument12 pagesResearch Paper NS11021, A Novel Opener of Large-Conductance Ca - Activated K Channels, Enhances Erectile Responses in RatsCarlos LabradaNo ratings yet
- Vol-1, Issue 7 Paper (5) Page 26-31Document7 pagesVol-1, Issue 7 Paper (5) Page 26-31IJMSHCNo ratings yet
- Acido Ursolico NNDocument5 pagesAcido Ursolico NNDIANA PATRICIA MOLANO PEREZNo ratings yet
- Volatile Anaesthetic Myocardial Protection: A Review of The Current LiteratureDocument5 pagesVolatile Anaesthetic Myocardial Protection: A Review of The Current Literatureserena7205No ratings yet
- SSRN Id4401700Document94 pagesSSRN Id4401700Milan Petrik100% (1)
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDocument8 pagesAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshNo ratings yet
- Nai Et Al 2017 Protective Effect of Astaxanthin On Acute Cerebral Infarction in RatsDocument8 pagesNai Et Al 2017 Protective Effect of Astaxanthin On Acute Cerebral Infarction in RatsMennatallah AliNo ratings yet
- Efectos Del Ultrasonido en FibroblastosDocument7 pagesEfectos Del Ultrasonido en FibroblastosErika VillacisNo ratings yet
- Vasopressin in Septic ShockDocument8 pagesVasopressin in Septic ShockVaishnavi SheteNo ratings yet
- 1 s2.0 S0925443923001850 MainDocument11 pages1 s2.0 S0925443923001850 Mainviventius yoshuaNo ratings yet
- Quercetin HypertensiveDocument8 pagesQuercetin HypertensiveAprilihardini LaksmiNo ratings yet
- The Role of The NaDocument13 pagesThe Role of The NamrtiborhornyikNo ratings yet
- Toksik Liver 9Document13 pagesToksik Liver 9Mita SilviyaniNo ratings yet
- Georgescu2013 PDFDocument9 pagesGeorgescu2013 PDFILHAM 'K FOR ALLNo ratings yet
- Expresi PeriostinDocument5 pagesExpresi Periostinkhoirul imaNo ratings yet
- Wendy W. Wu, C. Savio Chan, D. James Surmeier and John F. DisterhoftDocument13 pagesWendy W. Wu, C. Savio Chan, D. James Surmeier and John F. DisterhoftFedrmNo ratings yet
- Effect of Rosiglitazone On RabbiDocument7 pagesEffect of Rosiglitazone On Rabbinasirasna899No ratings yet
- 2007-Effects and Mechanisms of Electroacupuncture at PC6 On Frequency of Transient Lower Esophageal Sphincter Relaxation in Cats PDFDocument8 pages2007-Effects and Mechanisms of Electroacupuncture at PC6 On Frequency of Transient Lower Esophageal Sphincter Relaxation in Cats PDFvictoregvNo ratings yet
- Involvement of Parasympathetic Pelvic Efferent Pathway in Psychological Stress-Induced DefecationDocument10 pagesInvolvement of Parasympathetic Pelvic Efferent Pathway in Psychological Stress-Induced DefecationHrvoje JakovacNo ratings yet
- Multi-scale Extracellular Matrix Mechanics and MechanobiologyFrom EverandMulti-scale Extracellular Matrix Mechanics and MechanobiologyYanhang ZhangNo ratings yet
- Embryology of The Heart and The Great VesselsDocument66 pagesEmbryology of The Heart and The Great VesselsNavya HegdeNo ratings yet
- Overhead Transparency Human AtlasDocument34 pagesOverhead Transparency Human Atlasdr tamerNo ratings yet
- Science - G9 - Week 2 (Lessons-4-6)Document46 pagesScience - G9 - Week 2 (Lessons-4-6)Rhyan Zero-four BaluyutNo ratings yet
- Through The Decades (Beta) - Blocker Use and Outcomes in Acute Coronary Syndromes.Document8 pagesThrough The Decades (Beta) - Blocker Use and Outcomes in Acute Coronary Syndromes.Tony Miguel Saba SabaNo ratings yet
- Epidemiology, Risk Factors, Pathogenesis, and Natural History of Abdominal Aortic Aneurysm - UpToDateDocument15 pagesEpidemiology, Risk Factors, Pathogenesis, and Natural History of Abdominal Aortic Aneurysm - UpToDateALVARO ARIASNo ratings yet
- Yarber Autopsy ReportDocument7 pagesYarber Autopsy ReportSarah BatchaNo ratings yet
- GEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Document202 pagesGEHC DICOM Conformance VividT8 T8Pro DOC1482248 Rev2Shantanu MandalNo ratings yet
- Falenissa - Pemicu 3 Blok Etika Dan Hukum KedokteranDocument108 pagesFalenissa - Pemicu 3 Blok Etika Dan Hukum KedokteranIcarus WingsNo ratings yet
- Logiq e Datasheet PDFDocument12 pagesLogiq e Datasheet PDFrca ieftinNo ratings yet
- Doppler Echocardiography For The Small Animal Practitioner June A Boon Full ChapterDocument67 pagesDoppler Echocardiography For The Small Animal Practitioner June A Boon Full Chapterelizabeth.myers417100% (12)
- 110 TOP SURGERY Multiple Choice Questions and Answers PDF - Medical Multiple Choice Questions PDFDocument11 pages110 TOP SURGERY Multiple Choice Questions and Answers PDF - Medical Multiple Choice Questions PDFaziz0% (1)
- BrahmaiahDocument395 pagesBrahmaiahBrahmaiah UpputuriNo ratings yet
- Anatomy of The AortaDocument19 pagesAnatomy of The AortaUma Sanggari ThirunavukarasuNo ratings yet
- Measurements in Radiology Made EasyDocument213 pagesMeasurements in Radiology Made EasySllavko K. KallfaNo ratings yet
- Full Download Test Bank For Human Embryology and Developmental Biology 6th Edition Carlson PDF Full ChapterDocument36 pagesFull Download Test Bank For Human Embryology and Developmental Biology 6th Edition Carlson PDF Full Chapterjuristvesicantg7egk100% (22)
- Histologi Organ Dan Sistem Sirkulasi PoltekkesDocument28 pagesHistologi Organ Dan Sistem Sirkulasi PoltekkesJeon WonwooNo ratings yet
- State Group CCMP MCQDocument28 pagesState Group CCMP MCQGirish WaruNo ratings yet
- Crochet Anatomical Human Heart: by Kyla RaaymakersDocument7 pagesCrochet Anatomical Human Heart: by Kyla RaaymakersJavier BejaranoNo ratings yet
- Anatomy Quest.Document11 pagesAnatomy Quest.Ade AlcarazNo ratings yet
- Anatomy Physiology of The BrainDocument107 pagesAnatomy Physiology of The BrainRadha RamineniNo ratings yet
- Acute Coronary Syndrome Case-Report: A U F Angeles City College of NursingDocument77 pagesAcute Coronary Syndrome Case-Report: A U F Angeles City College of NursingCarlo Joseph Intal LlacerNo ratings yet
- ITQR14Document6 pagesITQR14Vasu NallaluthanNo ratings yet
- Diaphragm AnatomyDocument19 pagesDiaphragm AnatomyasujithdrNo ratings yet
- Biology Mnemonics (Neet - Aiims - Jipmer)Document67 pagesBiology Mnemonics (Neet - Aiims - Jipmer)MyCollectionMtgNo ratings yet
- PDFDocument97 pagesPDFUmeshNo ratings yet
- Coronary Artery Disease - Case StudyDocument19 pagesCoronary Artery Disease - Case StudyJulieteeySarania100% (4)
- For Questions Number 1-5, Refer To Scenario BelowDocument48 pagesFor Questions Number 1-5, Refer To Scenario BelowPuravin Crishan VeerasamyNo ratings yet