Download as pdf or txt
You might also like
- Instant Download Ebook PDF Exploring Psychology 11th Edition by David Myers PDF ScribdDocument41 pagesInstant Download Ebook PDF Exploring Psychology 11th Edition by David Myers PDF Scribdfrank.kuan506100% (50)
- Vital Signs Checklist (Temperature BP PR RR)Document6 pagesVital Signs Checklist (Temperature BP PR RR)Shades ofyellow80% (5)
- Plant Secondary Metabolites of Antiviral PropertieDocument7 pagesPlant Secondary Metabolites of Antiviral PropertieSteve Vladimir Acedo LazoNo ratings yet
- Andreetal 2016 FrontplsciDocument18 pagesAndreetal 2016 Frontplscijose cipriani rendonNo ratings yet
- Cannabis Based Pesticides 2018Document13 pagesCannabis Based Pesticides 2018richard.schumacherNo ratings yet
- Isolation, Diversity, and Growth-Promoting Activities of Endophytic Bacteria From Tea Cultivars of Zijuan and Yunkang-10Document11 pagesIsolation, Diversity, and Growth-Promoting Activities of Endophytic Bacteria From Tea Cultivars of Zijuan and Yunkang-10maczweiNo ratings yet
- Antifungal Activity MuñaDocument14 pagesAntifungal Activity Muñavictor hugo olivares perezNo ratings yet
- Herbal Plants Research PaperDocument6 pagesHerbal Plants Research Papergmannevnd100% (1)
- Invitro Micropropagation of Piper Betle L.: Original Research ArticleDocument6 pagesInvitro Micropropagation of Piper Betle L.: Original Research ArticleRifky AlfeniNo ratings yet
- 3 KFX ZXR PPNK 9 H K84 ZKGFBCNDocument10 pages3 KFX ZXR PPNK 9 H K84 ZKGFBCNAlisson CirinoNo ratings yet
- The Problem: Derris Elliptica Benth, Locally RecognizedDocument26 pagesThe Problem: Derris Elliptica Benth, Locally RecognizedJan Angela BaylonNo ratings yet
- Author's Accepted Manuscript: Tithonia DiversifoliaDocument55 pagesAuthor's Accepted Manuscript: Tithonia DiversifoliaJorge Alejandro Noriega PimentelNo ratings yet
- All Terpene AGlimpseintotheBiosynthesisofTerpenoidsDocument19 pagesAll Terpene AGlimpseintotheBiosynthesisofTerpenoidsMENTARI_KAIROSNo ratings yet
- MANSOORI - Phytochem of L Camara Crude ExtractDocument14 pagesMANSOORI - Phytochem of L Camara Crude ExtractPatrick John Ancheta MariñasNo ratings yet
- SJGGT 2 111Document5 pagesSJGGT 2 111Blaxez YTNo ratings yet
- BotanyDocument28 pagesBotanySehar SirajNo ratings yet
- Lata Et Al (2016) in Vitro Mass Propagation of Cannabis Sativa L. A Protocolo Refinement Using Novel Aromatic CytokininDocument9 pagesLata Et Al (2016) in Vitro Mass Propagation of Cannabis Sativa L. A Protocolo Refinement Using Novel Aromatic CytokininJ. Joaquin Varela-RestrepoNo ratings yet
- Bioactive Compounds From Marine Derived Aspergillus Penicillium Talaromyces and Trichoderma Species Rosario Nicoletti (Editor)Document70 pagesBioactive Compounds From Marine Derived Aspergillus Penicillium Talaromyces and Trichoderma Species Rosario Nicoletti (Editor)peter.reed327100% (12)
- Studies On Biodiversity of Phylloplane and Endophytic Fungi From Mature Leaves of Medicinal Plant, Carica Papaya L. With Antimicrobial PotentialDocument6 pagesStudies On Biodiversity of Phylloplane and Endophytic Fungi From Mature Leaves of Medicinal Plant, Carica Papaya L. With Antimicrobial PotentialRanadive AnanthNo ratings yet
- Studies On Antimicrobial Activity of Medicinal Plants Against Human PathogensDocument5 pagesStudies On Antimicrobial Activity of Medicinal Plants Against Human PathogensAnonymous CwJeBCAXpNo ratings yet
- In Vitro Mass Propagation of Cannabis Sativa L. A Protocol RefinementDocument9 pagesIn Vitro Mass Propagation of Cannabis Sativa L. A Protocol RefinementFelipe VélezNo ratings yet
- Efecto de Extractos Alcoholicos de Ajo Allium SatiDocument16 pagesEfecto de Extractos Alcoholicos de Ajo Allium SatiBrian FuhrNo ratings yet
- Biocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaDocument9 pagesBiocontrol Potential of Endophytic Fungi in Medicinal Plants From Wuhan Botanical Garden in ChinaMaria BatoolNo ratings yet
- Paper 4Document7 pagesPaper 4Vardhana JanakiramanNo ratings yet
- 1 s2.0 S2352340919309059 MainDocument6 pages1 s2.0 S2352340919309059 MainESPINOSANo ratings yet
- Plantas CubanasDocument9 pagesPlantas CubanasJunior GonzalesNo ratings yet
- Antibacterial Screening of Aqueous Extracts of Some Medicinal Plant and Their Fractions Used As Antidiarrheal Agents in Kinshasa-Democratic Republic of CongoDocument20 pagesAntibacterial Screening of Aqueous Extracts of Some Medicinal Plant and Their Fractions Used As Antidiarrheal Agents in Kinshasa-Democratic Republic of CongoFrengkyNo ratings yet
- Alternanthera Philoxeroides (Mart.) GrisebDocument4 pagesAlternanthera Philoxeroides (Mart.) GrisebRigotti Br100% (1)
- FILE - 20210529 - 153336 - Naturally Occurring Anthocyanin Structure Functions and Biosyntheticpathway in Fruit Plants 2329 9029 1000187Document9 pagesFILE - 20210529 - 153336 - Naturally Occurring Anthocyanin Structure Functions and Biosyntheticpathway in Fruit Plants 2329 9029 1000187Nguyen Thi Thuy Duong B1606703No ratings yet
- BiorationalInsecticides 04Document40 pagesBiorationalInsecticides 04rehoboth nyamanjerereNo ratings yet
- 4 Ijasrapr20194Document8 pages4 Ijasrapr20194TJPRC PublicationsNo ratings yet
- Chemical Analysis and Antioxidant Activity of Essential Oils of Two Morphotypes of Lippia Alba Mill N E BR Ex Britton P Wilson VerbenaceaeDocument15 pagesChemical Analysis and Antioxidant Activity of Essential Oils of Two Morphotypes of Lippia Alba Mill N E BR Ex Britton P Wilson VerbenaceaeArchana JoshiNo ratings yet
- Antibacterial and Phytochemical Analysis of Ethnomedicinal PlantsDocument7 pagesAntibacterial and Phytochemical Analysis of Ethnomedicinal PlantsAnkit BhardwajNo ratings yet
- 2817 PDFDocument9 pages2817 PDFCaryl Ann C. SernadillaNo ratings yet
- Phytochemical Screening of Medicinal Plants ThesisDocument7 pagesPhytochemical Screening of Medicinal Plants Thesisrebeccaevansspringfield100% (2)
- Applied Bryology BryotechnologyDocument12 pagesApplied Bryology Bryotechnology-No ratings yet
- AjenjoDocument9 pagesAjenjovidalNo ratings yet
- Plants: Chemical Composition and Nematicidal Properties of Sixteen Essential Oils-A ReviewDocument12 pagesPlants: Chemical Composition and Nematicidal Properties of Sixteen Essential Oils-A ReviewSitsuna FsNo ratings yet
- 226 732 1 PBDocument12 pages226 732 1 PBAlbert FandohanNo ratings yet
- Phytochemical Screening and Antibacterial Properties of Henna (Lawsonia Inermis) ROOTS EXTRACTSDocument11 pagesPhytochemical Screening and Antibacterial Properties of Henna (Lawsonia Inermis) ROOTS EXTRACTSPutra PratamaNo ratings yet
- Antimicrobial Properties and Biotransforming Ability of Fungal Endophytes From Ficus Carica L MoraceaeDocument26 pagesAntimicrobial Properties and Biotransforming Ability of Fungal Endophytes From Ficus Carica L MoraceaesnoelipoolNo ratings yet
- Isolate Endophytic Fungi From Local Catharanthus Roseus and Analyze Their Extracellular Enzyme ActivitiesDocument7 pagesIsolate Endophytic Fungi From Local Catharanthus Roseus and Analyze Their Extracellular Enzyme ActivitiesIJAR JOURNALNo ratings yet
- Constantinidou53 BacterialcolonizationofthephyllosphereofnineteenplantspeciesandantimicrobialactivityoftheirleafsecondarymetabolitesagainstleafassociatedbacteriaDocument11 pagesConstantinidou53 BacterialcolonizationofthephyllosphereofnineteenplantspeciesandantimicrobialactivityoftheirleafsecondarymetabolitesagainstleafassociatedbacteriaindahdwirahNo ratings yet
- Biologicalactivitiesof Alliumsativumand ZingiberofficinaleextractsDocument18 pagesBiologicalactivitiesof Alliumsativumand ZingiberofficinaleextractsEdgar alanNo ratings yet
- Chapter 3Document16 pagesChapter 3olotu_olaseindeNo ratings yet
- Optimisation and Characterisation of The Orange Pigment Produced by A Cold Adapted Strain of Penicillium Sp. GBPI P155 Isolated From Mountain EcosysDocument13 pagesOptimisation and Characterisation of The Orange Pigment Produced by A Cold Adapted Strain of Penicillium Sp. GBPI P155 Isolated From Mountain EcosysYeison EspinalNo ratings yet
- Biopolym - Cell 2018 34 6 461 enDocument16 pagesBiopolym - Cell 2018 34 6 461 enАнна ШаповаловаNo ratings yet
- Statia 20Document9 pagesStatia 20nguyen minh hienNo ratings yet
- PhytochemicalDocument15 pagesPhytochemicalfatimaNo ratings yet
- Antimicrobial, Antioxidant Activity and Phytochemical Prospection of Eugenia Involucrata DC. Leaf ExtractsDocument9 pagesAntimicrobial, Antioxidant Activity and Phytochemical Prospection of Eugenia Involucrata DC. Leaf ExtractsRobert Pereira dos SantosNo ratings yet
- A Comprehensive Review and Evaluation of in Vitro Antifungal Potential of Rauvolfia SerpentinaDocument14 pagesA Comprehensive Review and Evaluation of in Vitro Antifungal Potential of Rauvolfia SerpentinaIJRASETPublicationsNo ratings yet
- 1 s2.0 S030881461630276X MainVaccinium Myrtillus L.Document9 pages1 s2.0 S030881461630276X MainVaccinium Myrtillus L.JokililaNo ratings yet
- I J of Pharma NDP Harm Science 2016Document8 pagesI J of Pharma NDP Harm Science 2016JOSEPHNo ratings yet
- Cannabis Phenolics and Their BioactivitiesDocument44 pagesCannabis Phenolics and Their BioactivitiesTomislav ĆurićNo ratings yet
- Ultrasound-Assisted Extraction of Cannabinoids From Cannabis Sativa L. Optimized by Response Surface Methodology: Extraction of CannabinoidsDocument12 pagesUltrasound-Assisted Extraction of Cannabinoids From Cannabis Sativa L. Optimized by Response Surface Methodology: Extraction of CannabinoidsLabconquim SAS LaboratorioNo ratings yet
- Phytochemical and Antibacterial Activity Screening of Acalypha Indica and Ocimum TenuiflorumDocument9 pagesPhytochemical and Antibacterial Activity Screening of Acalypha Indica and Ocimum TenuiflorumIJRASETPublicationsNo ratings yet
- Trends in Herbal Usage - A Survey Study: Alexieva, I.N., Popova, A.T. and Mihaylova, D.SPDocument7 pagesTrends in Herbal Usage - A Survey Study: Alexieva, I.N., Popova, A.T. and Mihaylova, D.SPalibabaoleolaNo ratings yet
- Secondary MetabolitesDocument12 pagesSecondary MetabolitesJose Luis Perez MirandaNo ratings yet
- 2ARGDocument7 pages2ARGEmmanuel ucheNo ratings yet
- DiatomeasDocument20 pagesDiatomeasVictoria CruzNo ratings yet
- Diversity of Leaf Surface Fungi On The Medicinal Plant, Plectranthus Amboinicus Lour. and Its Antimicrobial Efficacy Against MTCC PathogensDocument6 pagesDiversity of Leaf Surface Fungi On The Medicinal Plant, Plectranthus Amboinicus Lour. and Its Antimicrobial Efficacy Against MTCC PathogensRanadive AnanthNo ratings yet
- Principles of Musculoskeletal Assessment: Introduction To Clinical Studies Traumatology RHS 231 Dr. Einas Al-EisaDocument36 pagesPrinciples of Musculoskeletal Assessment: Introduction To Clinical Studies Traumatology RHS 231 Dr. Einas Al-EisaroshinisureshNo ratings yet
- The Skeletal System - Multiple-Choice Questions PDFDocument4 pagesThe Skeletal System - Multiple-Choice Questions PDFJemeraldNo ratings yet
- Cell Biology Thesis TopicsDocument6 pagesCell Biology Thesis Topicsyeuroqaeg100% (2)
- EntomologyDocument32 pagesEntomologyAnnie Aragona-SatoNo ratings yet
- Regulation of Glycogenesis and GlyogenolysisDocument5 pagesRegulation of Glycogenesis and GlyogenolysismitalNo ratings yet
- Anatomy and Histology of The Normal Lung and AirwaysDocument20 pagesAnatomy and Histology of The Normal Lung and AirwaysFahd Abdullah Al-refaiNo ratings yet
- PDF Beta Oxid of FatDocument13 pagesPDF Beta Oxid of Fatshivani.priyaNo ratings yet
- Deal Siklus SelDocument50 pagesDeal Siklus Selanisa rahmaNo ratings yet
- Medicine Lecture 17,18Document51 pagesMedicine Lecture 17,18Nayela AkramNo ratings yet
- Slide Materi Dr. Nelfi, SPKFR - Osteoporosis and Fracture Prevention - PMR4GP 2022Document32 pagesSlide Materi Dr. Nelfi, SPKFR - Osteoporosis and Fracture Prevention - PMR4GP 2022Maulia Wisda Era ChresiaNo ratings yet
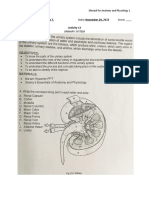
- Urinary SystemDocument5 pagesUrinary SystemJushelle Anne Tigoy Pilare100% (1)
- SciolyDocument91 pagesSciolyaarnavskrishnaNo ratings yet
- 8-Drugs Used in Anxiety and Panic Disprder (Edited)Document11 pages8-Drugs Used in Anxiety and Panic Disprder (Edited)ana01210761809No ratings yet
- Summative 7Document4 pagesSummative 7SPEILBERG LUMBAYNo ratings yet
- 1877-Article Text-1873-1-10-20080129Document18 pages1877-Article Text-1873-1-10-20080129Andreas KarasNo ratings yet
- Combined Science Notes-1Document258 pagesCombined Science Notes-1Premier Car Sales67% (3)
- Experiments On Geotropism An Experiment To Demonstrate That The Root Is Positively Geotropic (Or The Root Shows Positive Geotropism) Apparatus or MaterialsDocument4 pagesExperiments On Geotropism An Experiment To Demonstrate That The Root Is Positively Geotropic (Or The Root Shows Positive Geotropism) Apparatus or MaterialsMutegeki AdrianNo ratings yet
- Micro CH 2a StructureDocument155 pagesMicro CH 2a StructureBernadette Joyce PascualNo ratings yet
- Cartilage: Ethel Marie M. Mangada, RMTDocument34 pagesCartilage: Ethel Marie M. Mangada, RMTAesthetics MinNo ratings yet
- Cannabis Use and Cannabis Use DisorderDocument24 pagesCannabis Use and Cannabis Use DisorderHector AngelNo ratings yet
- 3.1 Answers: RememberingDocument13 pages3.1 Answers: RememberingDanny TNo ratings yet
- The Internal and External Anatomy of FishDocument5 pagesThe Internal and External Anatomy of FishGienelle BermidoNo ratings yet
- Pediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and ManagementDocument12 pagesPediatric Traumatic Brain Injury: Characteristic Features, Diagnosis, and ManagementsylvetriNo ratings yet
- In2Med HaematologyDocument30 pagesIn2Med Haematologye148659No ratings yet
- Unit:-II Chapter-6. Animal Tissue: Important PointsDocument11 pagesUnit:-II Chapter-6. Animal Tissue: Important PointsChandan KumarNo ratings yet
- Prelim Module in General PsychologyDocument9 pagesPrelim Module in General PsychologyAllona Zyra CambroneroNo ratings yet
- Question PaperDocument19 pagesQuestion PaperSaad NadafNo ratings yet
- Fungi and Allied Microbes - (10.9 Pilobolus)Document2 pagesFungi and Allied Microbes - (10.9 Pilobolus)vivek usNo ratings yet