
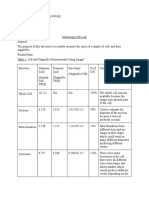
Tspiralis Rev
Tspiralis Rev
You might also like
- Aminoffs Electrodiagnosis in Clinical NeurologyDocument851 pagesAminoffs Electrodiagnosis in Clinical NeurologyMarysol Ulloa100% (1)
- OceanofPDF - Com Ultraviolet - Yvonne NavarroDocument213 pagesOceanofPDF - Com Ultraviolet - Yvonne Navarrobumble blick crumplestickNo ratings yet
- Measuring Cells LabDocument3 pagesMeasuring Cells LabClaudia100% (2)
- Prion NormalDocument6 pagesPrion NormalLisette Velásquez AnticonaNo ratings yet
- Art. 1. Agamemnon 1999Document5 pagesArt. 1. Agamemnon 1999Nara Elda Flores GonzalezNo ratings yet
- Ernfors P, Kucera J, Lee KF, Et Al. (1995)Document9 pagesErnfors P, Kucera J, Lee KF, Et Al. (1995)Rusda AnandaNo ratings yet
- Faria2006 PDFDocument11 pagesFaria2006 PDFPaige MunroeNo ratings yet
- M. Wintzer Et Al - Strategies For Identifying Genes That Play A Role in Spinal Cord RegenerationDocument9 pagesM. Wintzer Et Al - Strategies For Identifying Genes That Play A Role in Spinal Cord RegenerationHutsDMNo ratings yet
- Bangs 2000Document12 pagesBangs 2000laciyeg352No ratings yet
- Endogenous Retrovirus Sequences and Their UsefulneDocument1 pageEndogenous Retrovirus Sequences and Their Usefulnex994thng65No ratings yet
- Directing Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Document6 pagesDirecting Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Rebeca ChávezNo ratings yet
- Thompson1995 ScienceDocument8 pagesThompson1995 SciencePilar AufrastoNo ratings yet
- Currie, 2016 PDFDocument18 pagesCurrie, 2016 PDF22gaefNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- Translating The Histone Code: Science September 2001Document8 pagesTranslating The Histone Code: Science September 2001bra ritNo ratings yet
- Stem Cells: Dr. Valerie Bracchi-RicardDocument49 pagesStem Cells: Dr. Valerie Bracchi-RicardE12H12No ratings yet
- Des Car GableDocument10 pagesDes Car GableDago CastilloNo ratings yet
- 2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusDocument8 pages2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusNityananda PortelladaNo ratings yet
- Animal Models 1Document4 pagesAnimal Models 1Sanem CimenNo ratings yet
- 10 FullDocument9 pages10 FullDokdem AjaNo ratings yet
- Pezzotti 2014Document15 pagesPezzotti 2014maria.pezzottiNo ratings yet
- Marchioro Et Al. - 2005 - Postnatal Neurogenesis in The Medial Cortex of The Tropical Lizard Tropidurus HispidusDocument7 pagesMarchioro Et Al. - 2005 - Postnatal Neurogenesis in The Medial Cortex of The Tropical Lizard Tropidurus HispidusOsny SillasNo ratings yet
- Neural Stem Cells: Developmental Mechanisms and Disease ModelingDocument6 pagesNeural Stem Cells: Developmental Mechanisms and Disease ModelingDi RaNo ratings yet
- Caspases: Enemies WithinDocument6 pagesCaspases: Enemies WithinJuan manuel jiménez estradaNo ratings yet
- Neurogeneisis in RatsDocument7 pagesNeurogeneisis in RatsjsaccuzzoNo ratings yet
- The Neural Mechanisms of Gustation: A Distributed Processing CodeDocument12 pagesThe Neural Mechanisms of Gustation: A Distributed Processing CodesylNo ratings yet
- Hariri Et AlDocument4 pagesHariri Et AlJenniffer RiveraNo ratings yet
- Schlagg Ar 1991Document5 pagesSchlagg Ar 1991LeFührer Zabdiel PazaranNo ratings yet
- A Neuron's PerspectiveDocument15 pagesA Neuron's PerspectivePiali ChatterjeeNo ratings yet
- Genes Synap Atten MSDocument6 pagesGenes Synap Atten MSjohn fossellaNo ratings yet
- Micro ArrayDocument5 pagesMicro Arrayapi-3700537No ratings yet
- 2019 No PIIS1550413119300701Document3 pages2019 No PIIS1550413119300701nargess1981No ratings yet
- Cap 1 Parasitic NematodeDocument24 pagesCap 1 Parasitic NematodeJorge Chavez FloresNo ratings yet
- 115-Article Text-440-1-10-20180423Document10 pages115-Article Text-440-1-10-20180423Bronwyn Alayne H-lNo ratings yet
- 1642 FullDocument18 pages1642 FullFahrunnisa NurdinNo ratings yet
- Xenopus, An Emerging Model For Studying Pathologies of The Neural CrestDocument36 pagesXenopus, An Emerging Model For Studying Pathologies of The Neural CrestAIDAN AsigraNo ratings yet
- International Journal For ParasitologyDocument10 pagesInternational Journal For ParasitologyGabriel AvilaNo ratings yet
- Basic Concepts of EpigeneticsDocument10 pagesBasic Concepts of EpigeneticsRaju MehtaNo ratings yet
- Lammie Et Al 1994 Expression of C Kit and Kit Ligand Proteins in Normal Human TissuesDocument9 pagesLammie Et Al 1994 Expression of C Kit and Kit Ligand Proteins in Normal Human Tissuesvcox4460No ratings yet
- Lavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexDocument8 pagesLavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexCris AdFNo ratings yet
- Ramos Et Al 2007 DefinitivoDocument9 pagesRamos Et Al 2007 Definitivolucasmonje1978No ratings yet
- Perspective: Age-Related Decline in Neurogenesis: Old Cells or Old Environment?Document6 pagesPerspective: Age-Related Decline in Neurogenesis: Old Cells or Old Environment?Fahmi IsmailNo ratings yet
- Shirazi Et Al 2015 - Vitamin D and AstrocytesDocument15 pagesShirazi Et Al 2015 - Vitamin D and Astrocytesdr. RiyanNo ratings yet
- Preterm Labor - One Syndrome, Many CausesDocument7 pagesPreterm Labor - One Syndrome, Many CausesHaritza AqilahNo ratings yet
- Inventar Memoria en El HipocampusDocument5 pagesInventar Memoria en El HipocampuscorazondelatorNo ratings yet
- Human Motor Neuron Obtained From iPSCS As The Most Promising Tool To Define Pathomechanismsand Novel Therapies in ALSDocument2 pagesHuman Motor Neuron Obtained From iPSCS As The Most Promising Tool To Define Pathomechanismsand Novel Therapies in ALSRemedy PublicationsNo ratings yet
- Kitao 1996Document9 pagesKitao 1996M4shroomNo ratings yet
- 1.1 Article (Dragged) 7Document1 page1.1 Article (Dragged) 7OIJAsdiaNo ratings yet
- Walsh 2000 PDFDocument7 pagesWalsh 2000 PDFChrysoula GkaniNo ratings yet
- 1183 FullDocument5 pages1183 FullVijendra KavatalkarNo ratings yet
- Conventional and Non-Conventional Drosophila Toll Signaling 2Document20 pagesConventional and Non-Conventional Drosophila Toll Signaling 2Christian PakadangNo ratings yet
- p75 Neurotrophin Receptor Expression Defines A Population of BDNF-Responsive Neurogenic Precursor CellsDocument10 pagesp75 Neurotrophin Receptor Expression Defines A Population of BDNF-Responsive Neurogenic Precursor Cellsmarej312No ratings yet
- Jurnal HomeostatisDocument13 pagesJurnal HomeostatisJastip YueNo ratings yet
- Current Biology Vol 20 No 3 R108Document3 pagesCurrent Biology Vol 20 No 3 R108api-26133294No ratings yet
- 5437 FullDocument14 pages5437 FullFahrunnisa NurdinNo ratings yet
- Silence of The ROSDocument3 pagesSilence of The ROSEnzoNo ratings yet
- Perceptual-Learning 2017 Current-BiologyDocument6 pagesPerceptual-Learning 2017 Current-BiologyRolando Sepúlveda GonzálezNo ratings yet
- Nihms 144952Document9 pagesNihms 144952angelos96papNo ratings yet
- What Have Organoids and Assembloids Taught Us About The Pat - 2023 - BiologicalDocument10 pagesWhat Have Organoids and Assembloids Taught Us About The Pat - 2023 - Biologicalg.colleenNo ratings yet
- Dietary Restriction Enhances Neurotrophin Expression and Neurogenesis in The Hippocampus of Adult MiceDocument9 pagesDietary Restriction Enhances Neurotrophin Expression and Neurogenesis in The Hippocampus of Adult MiceJean Pierre Chastre LuzaNo ratings yet
- Transforming Growth Factor-A1 Is A Negative Modulator of Adult NeurogenesisDocument13 pagesTransforming Growth Factor-A1 Is A Negative Modulator of Adult Neurogenesisdr. RiyanNo ratings yet
- Spring '23 - Makakilo NPDocument4 pagesSpring '23 - Makakilo NPbumble blick crumplestickNo ratings yet
- Magic MoonDocument2 pagesMagic Moonbumble blick crumplestick0% (1)
- URLLinkDocument1 pageURLLinkbumble blick crumplestickNo ratings yet
- Acsnano 0c00913Document9 pagesAcsnano 0c00913bumble blick crumplestickNo ratings yet
- How Well Do You Like YourselfDocument2 pagesHow Well Do You Like Yourselfbumble blick crumplestickNo ratings yet
- Basics of Order and ChaosDocument2 pagesBasics of Order and Chaosbumble blick crumplestickNo ratings yet
- Rolaf EDITEDDocument4 pagesRolaf EDITEDbumble blick crumplestickNo ratings yet
- Toepke 2007Document5 pagesToepke 2007bumble blick crumplestickNo ratings yet
- Corpus Malicious Demo BookletDocument61 pagesCorpus Malicious Demo Bookletbumble blick crumplestickNo ratings yet
- Evil CampaignDocument1 pageEvil Campaignbumble blick crumplestickNo ratings yet
- Let The Galaxy BurnDocument165 pagesLet The Galaxy Burnbumble blick crumplestick0% (1)
- Avarice Daughter of Levistus: STR 10 (+0) DEX 18 (+4) CON 14 (+2) INT 20 (+5) WIS 10 (+0) Cha 18 (+4)Document3 pagesAvarice Daughter of Levistus: STR 10 (+0) DEX 18 (+4) CON 14 (+2) INT 20 (+5) WIS 10 (+0) Cha 18 (+4)bumble blick crumplestickNo ratings yet
- 51 Practice Question For Upcoming Pgimer MSC Nursing Entrance ExamDocument3 pages51 Practice Question For Upcoming Pgimer MSC Nursing Entrance ExamVrindha VijayanNo ratings yet
- MeltdownDocument92 pagesMeltdownMyles SamNo ratings yet
- Maglaqui - Skeletal SystemDocument5 pagesMaglaqui - Skeletal SystemNICOLE MAGLAQUINo ratings yet
- Assessment Nursing Diagnosis Planning Intervention Rationale Core CompetenciesDocument13 pagesAssessment Nursing Diagnosis Planning Intervention Rationale Core CompetenciesMina RacadioNo ratings yet
- Basic Life Support: Green Cross TrainingDocument56 pagesBasic Life Support: Green Cross TrainingJordanBangotNo ratings yet
- Topical Nitric Oxide Local Androgen Therapy PDFDocument6 pagesTopical Nitric Oxide Local Androgen Therapy PDFLuigi PolitoNo ratings yet
- Blood & Biologicals Section 2Document33 pagesBlood & Biologicals Section 2Louise Emy ViolettaNo ratings yet
- Fluid Volume Disturbances 2Document32 pagesFluid Volume Disturbances 2coosa liquorsNo ratings yet
- SmofKabiven Emulsion For Infusion ENG SMPC - 09001be6801b2eb8Document11 pagesSmofKabiven Emulsion For Infusion ENG SMPC - 09001be6801b2eb8Kirsten HartmanNo ratings yet
- Study Plan - 04 - Class 11 - PCMBDocument12 pagesStudy Plan - 04 - Class 11 - PCMBPranjalNo ratings yet
- Neonatal Ventilation, Step by StepDocument99 pagesNeonatal Ventilation, Step by StepMarely Ovando Castillo93% (14)
- Guide Selection Assessing SuitabilityDocument238 pagesGuide Selection Assessing SuitabilityMohamed Elmasry100% (1)
- Gaskell - Books From The Library of Walter Pagel - Part IIDocument180 pagesGaskell - Books From The Library of Walter Pagel - Part IICésar Cristov León Ortega100% (2)
- NewbornDocument2 pagesNewbornonijinoNo ratings yet
- Smith Dysmenorrhea and Menorrhagia - A Clinician's Guide PDFDocument163 pagesSmith Dysmenorrhea and Menorrhagia - A Clinician's Guide PDFsafiraNo ratings yet
- Malcolm Arnold Hurdling MasterclassDocument167 pagesMalcolm Arnold Hurdling MasterclassChris RawlinsonNo ratings yet
- Castro HandoutsDocument12 pagesCastro HandoutslghmshariNo ratings yet
- A Subdural HematomaDocument12 pagesA Subdural HematomaGina Irene IshakNo ratings yet
- What's TCMDocument35 pagesWhat's TCMirzal100% (1)
- Physical Properties of Skin and Its ApplicationsDocument94 pagesPhysical Properties of Skin and Its ApplicationsdrshilpibhadaniNo ratings yet
- Mesotherapy RejuvenationDocument40 pagesMesotherapy RejuvenationerlanggawidyaputriNo ratings yet
- Dr. Dinesh Kumar Yadav, KDCHDocument56 pagesDr. Dinesh Kumar Yadav, KDCHDinesh Kr. YadavNo ratings yet
- Bio 407 - Basic EntomologyDocument79 pagesBio 407 - Basic Entomologyzombitrashh100% (1)
- 382 Thyroid Gland Physiology and TestingDocument7 pages382 Thyroid Gland Physiology and TestingJetmoon BlackNo ratings yet
- Pulpal IrritantsDocument10 pagesPulpal IrritantsDiverse AnunturiNo ratings yet
- Praktikum BiokimiaDocument148 pagesPraktikum BiokimiaZuniva AndanNo ratings yet
- Aula 4 - Eritrocitos MorfologiaDocument9 pagesAula 4 - Eritrocitos MorfologiarodrigomurielNo ratings yet
- Biological Control Systems: Biomedical Engineering - Bcs - Short Questions and AnswersDocument15 pagesBiological Control Systems: Biomedical Engineering - Bcs - Short Questions and AnswersNoor AhmedNo ratings yet



































































































Download as pdf or txt
You might also like
- Aminoffs Electrodiagnosis in Clinical NeurologyDocument851 pagesAminoffs Electrodiagnosis in Clinical NeurologyMarysol Ulloa100% (1)
- OceanofPDF - Com Ultraviolet - Yvonne NavarroDocument213 pagesOceanofPDF - Com Ultraviolet - Yvonne Navarrobumble blick crumplestickNo ratings yet
- Measuring Cells LabDocument3 pagesMeasuring Cells LabClaudia100% (2)
- Prion NormalDocument6 pagesPrion NormalLisette Velásquez AnticonaNo ratings yet
- Art. 1. Agamemnon 1999Document5 pagesArt. 1. Agamemnon 1999Nara Elda Flores GonzalezNo ratings yet
- Ernfors P, Kucera J, Lee KF, Et Al. (1995)Document9 pagesErnfors P, Kucera J, Lee KF, Et Al. (1995)Rusda AnandaNo ratings yet
- Faria2006 PDFDocument11 pagesFaria2006 PDFPaige MunroeNo ratings yet
- M. Wintzer Et Al - Strategies For Identifying Genes That Play A Role in Spinal Cord RegenerationDocument9 pagesM. Wintzer Et Al - Strategies For Identifying Genes That Play A Role in Spinal Cord RegenerationHutsDMNo ratings yet
- Bangs 2000Document12 pagesBangs 2000laciyeg352No ratings yet
- Endogenous Retrovirus Sequences and Their UsefulneDocument1 pageEndogenous Retrovirus Sequences and Their Usefulnex994thng65No ratings yet
- Directing Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Document6 pagesDirecting Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Rebeca ChávezNo ratings yet
- Thompson1995 ScienceDocument8 pagesThompson1995 SciencePilar AufrastoNo ratings yet
- Currie, 2016 PDFDocument18 pagesCurrie, 2016 PDF22gaefNo ratings yet
- Paper Canty 2009 - PV HolesDocument11 pagesPaper Canty 2009 - PV Holesmaariamunoz9No ratings yet
- Translating The Histone Code: Science September 2001Document8 pagesTranslating The Histone Code: Science September 2001bra ritNo ratings yet
- Stem Cells: Dr. Valerie Bracchi-RicardDocument49 pagesStem Cells: Dr. Valerie Bracchi-RicardE12H12No ratings yet
- Des Car GableDocument10 pagesDes Car GableDago CastilloNo ratings yet
- 2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusDocument8 pages2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusNityananda PortelladaNo ratings yet
- Animal Models 1Document4 pagesAnimal Models 1Sanem CimenNo ratings yet
- 10 FullDocument9 pages10 FullDokdem AjaNo ratings yet
- Pezzotti 2014Document15 pagesPezzotti 2014maria.pezzottiNo ratings yet
- Marchioro Et Al. - 2005 - Postnatal Neurogenesis in The Medial Cortex of The Tropical Lizard Tropidurus HispidusDocument7 pagesMarchioro Et Al. - 2005 - Postnatal Neurogenesis in The Medial Cortex of The Tropical Lizard Tropidurus HispidusOsny SillasNo ratings yet
- Neural Stem Cells: Developmental Mechanisms and Disease ModelingDocument6 pagesNeural Stem Cells: Developmental Mechanisms and Disease ModelingDi RaNo ratings yet
- Caspases: Enemies WithinDocument6 pagesCaspases: Enemies WithinJuan manuel jiménez estradaNo ratings yet
- Neurogeneisis in RatsDocument7 pagesNeurogeneisis in RatsjsaccuzzoNo ratings yet
- The Neural Mechanisms of Gustation: A Distributed Processing CodeDocument12 pagesThe Neural Mechanisms of Gustation: A Distributed Processing CodesylNo ratings yet
- Hariri Et AlDocument4 pagesHariri Et AlJenniffer RiveraNo ratings yet
- Schlagg Ar 1991Document5 pagesSchlagg Ar 1991LeFührer Zabdiel PazaranNo ratings yet
- A Neuron's PerspectiveDocument15 pagesA Neuron's PerspectivePiali ChatterjeeNo ratings yet
- Genes Synap Atten MSDocument6 pagesGenes Synap Atten MSjohn fossellaNo ratings yet
- Micro ArrayDocument5 pagesMicro Arrayapi-3700537No ratings yet
- 2019 No PIIS1550413119300701Document3 pages2019 No PIIS1550413119300701nargess1981No ratings yet
- Cap 1 Parasitic NematodeDocument24 pagesCap 1 Parasitic NematodeJorge Chavez FloresNo ratings yet
- 115-Article Text-440-1-10-20180423Document10 pages115-Article Text-440-1-10-20180423Bronwyn Alayne H-lNo ratings yet
- 1642 FullDocument18 pages1642 FullFahrunnisa NurdinNo ratings yet
- Xenopus, An Emerging Model For Studying Pathologies of The Neural CrestDocument36 pagesXenopus, An Emerging Model For Studying Pathologies of The Neural CrestAIDAN AsigraNo ratings yet
- International Journal For ParasitologyDocument10 pagesInternational Journal For ParasitologyGabriel AvilaNo ratings yet
- Basic Concepts of EpigeneticsDocument10 pagesBasic Concepts of EpigeneticsRaju MehtaNo ratings yet
- Lammie Et Al 1994 Expression of C Kit and Kit Ligand Proteins in Normal Human TissuesDocument9 pagesLammie Et Al 1994 Expression of C Kit and Kit Ligand Proteins in Normal Human Tissuesvcox4460No ratings yet
- Lavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexDocument8 pagesLavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexCris AdFNo ratings yet
- Ramos Et Al 2007 DefinitivoDocument9 pagesRamos Et Al 2007 Definitivolucasmonje1978No ratings yet
- Perspective: Age-Related Decline in Neurogenesis: Old Cells or Old Environment?Document6 pagesPerspective: Age-Related Decline in Neurogenesis: Old Cells or Old Environment?Fahmi IsmailNo ratings yet
- Shirazi Et Al 2015 - Vitamin D and AstrocytesDocument15 pagesShirazi Et Al 2015 - Vitamin D and Astrocytesdr. RiyanNo ratings yet
- Preterm Labor - One Syndrome, Many CausesDocument7 pagesPreterm Labor - One Syndrome, Many CausesHaritza AqilahNo ratings yet
- Inventar Memoria en El HipocampusDocument5 pagesInventar Memoria en El HipocampuscorazondelatorNo ratings yet
- Human Motor Neuron Obtained From iPSCS As The Most Promising Tool To Define Pathomechanismsand Novel Therapies in ALSDocument2 pagesHuman Motor Neuron Obtained From iPSCS As The Most Promising Tool To Define Pathomechanismsand Novel Therapies in ALSRemedy PublicationsNo ratings yet
- Kitao 1996Document9 pagesKitao 1996M4shroomNo ratings yet
- 1.1 Article (Dragged) 7Document1 page1.1 Article (Dragged) 7OIJAsdiaNo ratings yet
- Walsh 2000 PDFDocument7 pagesWalsh 2000 PDFChrysoula GkaniNo ratings yet
- 1183 FullDocument5 pages1183 FullVijendra KavatalkarNo ratings yet
- Conventional and Non-Conventional Drosophila Toll Signaling 2Document20 pagesConventional and Non-Conventional Drosophila Toll Signaling 2Christian PakadangNo ratings yet
- p75 Neurotrophin Receptor Expression Defines A Population of BDNF-Responsive Neurogenic Precursor CellsDocument10 pagesp75 Neurotrophin Receptor Expression Defines A Population of BDNF-Responsive Neurogenic Precursor Cellsmarej312No ratings yet
- Jurnal HomeostatisDocument13 pagesJurnal HomeostatisJastip YueNo ratings yet
- Current Biology Vol 20 No 3 R108Document3 pagesCurrent Biology Vol 20 No 3 R108api-26133294No ratings yet
- 5437 FullDocument14 pages5437 FullFahrunnisa NurdinNo ratings yet
- Silence of The ROSDocument3 pagesSilence of The ROSEnzoNo ratings yet
- Perceptual-Learning 2017 Current-BiologyDocument6 pagesPerceptual-Learning 2017 Current-BiologyRolando Sepúlveda GonzálezNo ratings yet
- Nihms 144952Document9 pagesNihms 144952angelos96papNo ratings yet
- What Have Organoids and Assembloids Taught Us About The Pat - 2023 - BiologicalDocument10 pagesWhat Have Organoids and Assembloids Taught Us About The Pat - 2023 - Biologicalg.colleenNo ratings yet
- Dietary Restriction Enhances Neurotrophin Expression and Neurogenesis in The Hippocampus of Adult MiceDocument9 pagesDietary Restriction Enhances Neurotrophin Expression and Neurogenesis in The Hippocampus of Adult MiceJean Pierre Chastre LuzaNo ratings yet
- Transforming Growth Factor-A1 Is A Negative Modulator of Adult NeurogenesisDocument13 pagesTransforming Growth Factor-A1 Is A Negative Modulator of Adult Neurogenesisdr. RiyanNo ratings yet
- Spring '23 - Makakilo NPDocument4 pagesSpring '23 - Makakilo NPbumble blick crumplestickNo ratings yet
- Magic MoonDocument2 pagesMagic Moonbumble blick crumplestick0% (1)
- URLLinkDocument1 pageURLLinkbumble blick crumplestickNo ratings yet
- Acsnano 0c00913Document9 pagesAcsnano 0c00913bumble blick crumplestickNo ratings yet
- How Well Do You Like YourselfDocument2 pagesHow Well Do You Like Yourselfbumble blick crumplestickNo ratings yet
- Basics of Order and ChaosDocument2 pagesBasics of Order and Chaosbumble blick crumplestickNo ratings yet
- Rolaf EDITEDDocument4 pagesRolaf EDITEDbumble blick crumplestickNo ratings yet
- Toepke 2007Document5 pagesToepke 2007bumble blick crumplestickNo ratings yet
- Corpus Malicious Demo BookletDocument61 pagesCorpus Malicious Demo Bookletbumble blick crumplestickNo ratings yet
- Evil CampaignDocument1 pageEvil Campaignbumble blick crumplestickNo ratings yet
- Let The Galaxy BurnDocument165 pagesLet The Galaxy Burnbumble blick crumplestick0% (1)
- Avarice Daughter of Levistus: STR 10 (+0) DEX 18 (+4) CON 14 (+2) INT 20 (+5) WIS 10 (+0) Cha 18 (+4)Document3 pagesAvarice Daughter of Levistus: STR 10 (+0) DEX 18 (+4) CON 14 (+2) INT 20 (+5) WIS 10 (+0) Cha 18 (+4)bumble blick crumplestickNo ratings yet
- 51 Practice Question For Upcoming Pgimer MSC Nursing Entrance ExamDocument3 pages51 Practice Question For Upcoming Pgimer MSC Nursing Entrance ExamVrindha VijayanNo ratings yet
- MeltdownDocument92 pagesMeltdownMyles SamNo ratings yet
- Maglaqui - Skeletal SystemDocument5 pagesMaglaqui - Skeletal SystemNICOLE MAGLAQUINo ratings yet
- Assessment Nursing Diagnosis Planning Intervention Rationale Core CompetenciesDocument13 pagesAssessment Nursing Diagnosis Planning Intervention Rationale Core CompetenciesMina RacadioNo ratings yet
- Basic Life Support: Green Cross TrainingDocument56 pagesBasic Life Support: Green Cross TrainingJordanBangotNo ratings yet
- Topical Nitric Oxide Local Androgen Therapy PDFDocument6 pagesTopical Nitric Oxide Local Androgen Therapy PDFLuigi PolitoNo ratings yet
- Blood & Biologicals Section 2Document33 pagesBlood & Biologicals Section 2Louise Emy ViolettaNo ratings yet
- Fluid Volume Disturbances 2Document32 pagesFluid Volume Disturbances 2coosa liquorsNo ratings yet
- SmofKabiven Emulsion For Infusion ENG SMPC - 09001be6801b2eb8Document11 pagesSmofKabiven Emulsion For Infusion ENG SMPC - 09001be6801b2eb8Kirsten HartmanNo ratings yet
- Study Plan - 04 - Class 11 - PCMBDocument12 pagesStudy Plan - 04 - Class 11 - PCMBPranjalNo ratings yet
- Neonatal Ventilation, Step by StepDocument99 pagesNeonatal Ventilation, Step by StepMarely Ovando Castillo93% (14)
- Guide Selection Assessing SuitabilityDocument238 pagesGuide Selection Assessing SuitabilityMohamed Elmasry100% (1)
- Gaskell - Books From The Library of Walter Pagel - Part IIDocument180 pagesGaskell - Books From The Library of Walter Pagel - Part IICésar Cristov León Ortega100% (2)
- NewbornDocument2 pagesNewbornonijinoNo ratings yet
- Smith Dysmenorrhea and Menorrhagia - A Clinician's Guide PDFDocument163 pagesSmith Dysmenorrhea and Menorrhagia - A Clinician's Guide PDFsafiraNo ratings yet
- Malcolm Arnold Hurdling MasterclassDocument167 pagesMalcolm Arnold Hurdling MasterclassChris RawlinsonNo ratings yet
- Castro HandoutsDocument12 pagesCastro HandoutslghmshariNo ratings yet
- A Subdural HematomaDocument12 pagesA Subdural HematomaGina Irene IshakNo ratings yet
- What's TCMDocument35 pagesWhat's TCMirzal100% (1)
- Physical Properties of Skin and Its ApplicationsDocument94 pagesPhysical Properties of Skin and Its ApplicationsdrshilpibhadaniNo ratings yet
- Mesotherapy RejuvenationDocument40 pagesMesotherapy RejuvenationerlanggawidyaputriNo ratings yet
- Dr. Dinesh Kumar Yadav, KDCHDocument56 pagesDr. Dinesh Kumar Yadav, KDCHDinesh Kr. YadavNo ratings yet
- Bio 407 - Basic EntomologyDocument79 pagesBio 407 - Basic Entomologyzombitrashh100% (1)
- 382 Thyroid Gland Physiology and TestingDocument7 pages382 Thyroid Gland Physiology and TestingJetmoon BlackNo ratings yet
- Pulpal IrritantsDocument10 pagesPulpal IrritantsDiverse AnunturiNo ratings yet
- Praktikum BiokimiaDocument148 pagesPraktikum BiokimiaZuniva AndanNo ratings yet
- Aula 4 - Eritrocitos MorfologiaDocument9 pagesAula 4 - Eritrocitos MorfologiarodrigomurielNo ratings yet
- Biological Control Systems: Biomedical Engineering - Bcs - Short Questions and AnswersDocument15 pagesBiological Control Systems: Biomedical Engineering - Bcs - Short Questions and AnswersNoor AhmedNo ratings yet