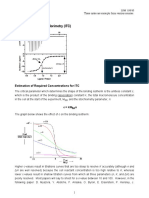
Download as pdf or txt
You might also like
- Isothermal Titration CalorimetryDocument6 pagesIsothermal Titration CalorimetryMiles NsgNo ratings yet
- Designing Itc v7 ExptsDocument5 pagesDesigning Itc v7 ExptsRigel_TNo ratings yet
- Modeling and Simulation of The Thermal Runaway in Cylindrical 18650 Lithium-Ion BatteriesDocument7 pagesModeling and Simulation of The Thermal Runaway in Cylindrical 18650 Lithium-Ion BatteriesKamel IbrahimNo ratings yet
- Itc Current ProtocolsDocument24 pagesItc Current ProtocolsRigel_TNo ratings yet
- Simple Calculations, Principles, and Techniques To Prevent Reactive Chemicals Events in The LaboratoryDocument4 pagesSimple Calculations, Principles, and Techniques To Prevent Reactive Chemicals Events in The LaboratorypiloNo ratings yet
- DNA Computing: A ReviewDocument12 pagesDNA Computing: A ReviewVinod SainiNo ratings yet
- IB HL Chemistry 5.1 Measuring Enthalpy ChangesDocument6 pagesIB HL Chemistry 5.1 Measuring Enthalpy ChangesAva PatriciaNo ratings yet
- Isothermal Titration Calorimetry: Presented By: Ms. Prajakta S.Pawar. Guided byDocument54 pagesIsothermal Titration Calorimetry: Presented By: Ms. Prajakta S.Pawar. Guided byVrushali Puranik-GokhaleNo ratings yet
- Unit 3 Textbook QuestionsDocument10 pagesUnit 3 Textbook QuestionsT MNo ratings yet
- Estudio y Descripcion de Mecanismos de Reaciones OrganicasDocument75 pagesEstudio y Descripcion de Mecanismos de Reaciones OrganicascarolinaNo ratings yet
- IEEE ACCESS Temperature Control For Newton's Law of Cooling Using PID ControllerDocument7 pagesIEEE ACCESS Temperature Control For Newton's Law of Cooling Using PID ControllerAlejandro MisesNo ratings yet
- Biophysical Chemistry: J. SeeligDocument7 pagesBiophysical Chemistry: J. SeeligLiv IabNo ratings yet
- Reading Modulatet DSC Termochimica Acta 1994Document13 pagesReading Modulatet DSC Termochimica Acta 1994pedroNo ratings yet
- Batch and Semibatch Reactor Performance For An Exothermic ReactionDocument8 pagesBatch and Semibatch Reactor Performance For An Exothermic ReactionPablo Solano Jr.No ratings yet
- Review of Theory of Dissipative AdaptationDocument3 pagesReview of Theory of Dissipative AdaptationGeorge HalalNo ratings yet
- J. Electrochem. Soc.-1995-Pals-3282-8Document7 pagesJ. Electrochem. Soc.-1995-Pals-3282-8Alhussein Mohammed Abd ElhafeezNo ratings yet
- Shivesh Sabbarwal, Prachi Srivastava, Vivek Kumar Verma, and Manoj KumarDocument17 pagesShivesh Sabbarwal, Prachi Srivastava, Vivek Kumar Verma, and Manoj KumarGogyNo ratings yet
- Grupo 8Document7 pagesGrupo 8YuneskyNo ratings yet
- Determination of A Reaction's Activated Energy Using Naoh As A ReactantDocument7 pagesDetermination of A Reaction's Activated Energy Using Naoh As A ReactantTops10JNo ratings yet
- Qua 83Document9 pagesQua 83jnmanivann21No ratings yet
- Mackintosh Paper PDFDocument4 pagesMackintosh Paper PDFheheNo ratings yet
- 1 s2.0 S2352152X20319587 MainDocument8 pages1 s2.0 S2352152X20319587 MainSatyajit PatilNo ratings yet
- Paper Eco SQ Abs Q 150502Document11 pagesPaper Eco SQ Abs Q 150502Laser AerospaceNo ratings yet
- 1 s2.0 S0378775321013549 MainDocument10 pages1 s2.0 S0378775321013549 MainArghya MukherjeeNo ratings yet
- Gibbs Paradox SolutionDocument11 pagesGibbs Paradox SolutioncraverNo ratings yet
- Ref 1Document4 pagesRef 1soroushNo ratings yet
- Physica E: Low-Dimensional Systems and Nanostructures: Yanchao Zhang, Jincan Chen TDocument4 pagesPhysica E: Low-Dimensional Systems and Nanostructures: Yanchao Zhang, Jincan Chen TAstrid Camila Riveros MesaNo ratings yet
- Thermodynamic Calculations For Biochemical Transport and Reaction Processes in Metabolic NetworksDocument9 pagesThermodynamic Calculations For Biochemical Transport and Reaction Processes in Metabolic NetworksDizaichaNo ratings yet
- BLT 05 227Document14 pagesBLT 05 227Galuh WiedaniNo ratings yet
- Thermodynamic Calculations For Biochemical Transport and Reaction Processes in Metabolic NetworksDocument9 pagesThermodynamic Calculations For Biochemical Transport and Reaction Processes in Metabolic NetworksDizaichaNo ratings yet
- Differences in The Energetics of Collagen Denaturation in ConnectiveDocument8 pagesDifferences in The Energetics of Collagen Denaturation in ConnectiveBowie IraNo ratings yet
- Composites: Part A: Oleksandr G. Kravchenko, Chunyu Li, Alejandro Strachan, Sergii G. Kravchenko, R. Byron PipesDocument9 pagesComposites: Part A: Oleksandr G. Kravchenko, Chunyu Li, Alejandro Strachan, Sergii G. Kravchenko, R. Byron Pipesmarco_ravelo_10No ratings yet
- Philip 1981Document9 pagesPhilip 1981swatiNo ratings yet
- CHM71 - Detailed KPI'sDocument3 pagesCHM71 - Detailed KPI'sFatima BinIshaqNo ratings yet
- Energies 06 02709Document17 pagesEnergies 06 02709Anonymous raLlP6WyNo ratings yet
- Chemistry 7th Edition Mcmurry Solutions ManualDocument13 pagesChemistry 7th Edition Mcmurry Solutions Manualkaitlynmosleyewigyrapof100% (40)
- Uncertainty of Measurements: Experiment 2Document2 pagesUncertainty of Measurements: Experiment 2Marjory CastilloNo ratings yet
- The Effect of Different Sky Temperature Models On The Accuracy in The Estimation of The Performance of A Photovoltaic ModuleDocument6 pagesThe Effect of Different Sky Temperature Models On The Accuracy in The Estimation of The Performance of A Photovoltaic ModuleJovanJoksicNo ratings yet
- Ebook Chemistry 7Th Edition Mcmurry Solutions Manual Full Chapter PDFDocument34 pagesEbook Chemistry 7Th Edition Mcmurry Solutions Manual Full Chapter PDFricinussquabash.46iz9100% (15)
- Analytical Chemistry (2006), 78 (8), 2684-2693.Document10 pagesAnalytical Chemistry (2006), 78 (8), 2684-2693.Leo G. La PlataNo ratings yet
- Relation Between The Cell Temperature of A HCPV Module and Atmospheric ParametersDocument6 pagesRelation Between The Cell Temperature of A HCPV Module and Atmospheric ParametersAvi NivNo ratings yet
- Exp 1 Calorimetry IntroDocument3 pagesExp 1 Calorimetry IntroMarco FranciscoNo ratings yet
- Application of Isothermal Titration Calorimetry in The Biological Sciences: Things Are Heating Up!Document3 pagesApplication of Isothermal Titration Calorimetry in The Biological Sciences: Things Are Heating Up!Jack HsuNo ratings yet
- Modeling and Analysis of Renewable Pem Fuel Cell System: SciencedirectDocument15 pagesModeling and Analysis of Renewable Pem Fuel Cell System: SciencedirectAnonymous 78iAn6No ratings yet
- Heat Transfer Correlations For Multilayer Insulation SystemsDocument5 pagesHeat Transfer Correlations For Multilayer Insulation SystemsPankil ShahNo ratings yet
- 1 s2.0 S187538921100678X Main PDFDocument7 pages1 s2.0 S187538921100678X Main PDFSohom ChakrabortyNo ratings yet
- General Chemistry: ThermochemistryDocument60 pagesGeneral Chemistry: ThermochemistryOrxan ƏhmədovNo ratings yet
- Heat-Transfer Correlations For Natural Convection BoilingDocument15 pagesHeat-Transfer Correlations For Natural Convection BoilingForcus on100% (1)
- Dielectric Frequency Response Measurements and Dissipation Factor Temperature DependenceDocument5 pagesDielectric Frequency Response Measurements and Dissipation Factor Temperature Dependencekong shiaw huiNo ratings yet
- Experimental Research of Heat Transfer in Cooling Tower: KeywordsDocument6 pagesExperimental Research of Heat Transfer in Cooling Tower: Keywordsashfaque4155No ratings yet
- Prater Vol 8 284 1958Document3 pagesPrater Vol 8 284 1958Dai DomNo ratings yet
- Numerical and Experimental Study On Thermal Behavior of Prismatic Lithium-Ion Battery For Large-Capacity Energy StorageDocument15 pagesNumerical and Experimental Study On Thermal Behavior of Prismatic Lithium-Ion Battery For Large-Capacity Energy StoragekienfinanceNo ratings yet
- Chapter 7 ThermochemistryDocument43 pagesChapter 7 ThermochemistryGörkem Damdere100% (1)
- Li2002 Article TheCoarseningKineticsOfΓParticDocument7 pagesLi2002 Article TheCoarseningKineticsOfΓPartickandula.munikumarNo ratings yet
- National Research Conseil National Council Canada de Recherches CanadaDocument33 pagesNational Research Conseil National Council Canada de Recherches Canadamanoj thankachanNo ratings yet
- Electrophoresis (2005), 26 (22), 4333-4344Document12 pagesElectrophoresis (2005), 26 (22), 4333-4344Leo G. La PlataNo ratings yet
- Energetics NoteDocument9 pagesEnergetics Notecupcakesen22No ratings yet
- A Modern Course in Statistical PhysicsFrom EverandA Modern Course in Statistical PhysicsRating: 3.5 out of 5 stars3.5/5 (2)
- Reviews in Computational ChemistryFrom EverandReviews in Computational ChemistryAbby L. ParrillNo ratings yet
- Entropy NotesDocument8 pagesEntropy NotesTadesse AyalewNo ratings yet
- Ramiro Augusto Salazar La Rotta, Ing. Químico, PH.D., CC: 91227727 Colombia (UIS, SENA) Juan Manuel Salazar Villamizar (Hijo) - E-Mail:, AstractDocument9 pagesRamiro Augusto Salazar La Rotta, Ing. Químico, PH.D., CC: 91227727 Colombia (UIS, SENA) Juan Manuel Salazar Villamizar (Hijo) - E-Mail:, AstractRamiro Augusto salazar La RottaNo ratings yet
- Health Effects of Subtle EnergiesDocument10 pagesHealth Effects of Subtle EnergiesSaharaNo ratings yet
- cHME CDFDocument15 pagescHME CDFSulochana KoviNo ratings yet
- UG Syllabus 2017 18Document53 pagesUG Syllabus 2017 18Bhuvanesh wari WariNo ratings yet
- Basic Thermodynamics (BTME-305-18)Document14 pagesBasic Thermodynamics (BTME-305-18)Surjit Kumar GandhiNo ratings yet
- Thermal 1 SyllabusDocument10 pagesThermal 1 SyllabussoundarinboxNo ratings yet
- Sample ActivityDocument4 pagesSample ActivityJeson PawaonNo ratings yet
- Definition of ThermodynamicsDocument8 pagesDefinition of ThermodynamicsDavid MahrezNo ratings yet
- Thermodynamic Exercise EntropyDocument2 pagesThermodynamic Exercise EntropyFarid AimanNo ratings yet
- TC and TD Paper 1Document1 pageTC and TD Paper 1Anant RajNo ratings yet
- Thermodynamics of Polymer SolDocument63 pagesThermodynamics of Polymer Solgopi kumar pramanikNo ratings yet
- William Eckhardt - Causal Time Asymmetry.131100210Document43 pagesWilliam Eckhardt - Causal Time Asymmetry.131100210SinoNo ratings yet
- Classical Theory of DetonationDocument20 pagesClassical Theory of Detonationalexander.lhoistNo ratings yet
- Lesson 1 ThermodynamicsDocument40 pagesLesson 1 ThermodynamicsRex OabelNo ratings yet
- Statistical Methods and Thermodynamics - BatistaDocument97 pagesStatistical Methods and Thermodynamics - BatistapepeperezNo ratings yet
- B Pharmacy 1Document64 pagesB Pharmacy 1api-221326191No ratings yet
- PHYS121ECE2Document19 pagesPHYS121ECE2Rebuyon SharbelNo ratings yet
- Chapter 17 KeyDocument16 pagesChapter 17 KeyAriny ZaqiyahNo ratings yet
- BSC Chemistry SyllabusDocument22 pagesBSC Chemistry Syllabusshivamrawat7557No ratings yet
- Witten - QM of A Black HoleDocument4 pagesWitten - QM of A Black HoleAbhijeet BhalkikarNo ratings yet
- Statistical Mechanics: Daniel F. Styer DecemberDocument258 pagesStatistical Mechanics: Daniel F. Styer DecemberTarun FreemanNo ratings yet
- Cryogenic PhysicsDocument27 pagesCryogenic PhysicsOmongo EmmanuelNo ratings yet
- Laidler Physical Chemistry 4th EditionDocument1,078 pagesLaidler Physical Chemistry 4th EditionqamaralmahseriNo ratings yet
- Physics Model PaperDocument63 pagesPhysics Model Paperapi-3699388100% (2)
- 2 4-TeDocument197 pages2 4-TeJohann Roy CruzNo ratings yet
- Industrial EngineeringDocument84 pagesIndustrial EngineeringmuetanianNo ratings yet
- Baecker, D. - A Note On Ludwig Von Bertalanffy and The Form Problem of LifeDocument10 pagesBaecker, D. - A Note On Ludwig Von Bertalanffy and The Form Problem of LifeJorge GonzálezNo ratings yet
- BSC - (Phy - Chem - Maths)Document74 pagesBSC - (Phy - Chem - Maths)Shravani SalunkheNo ratings yet
- List of Thermodynamic Properties - Wikipedia, The Free EncyclopediaDocument3 pagesList of Thermodynamic Properties - Wikipedia, The Free EncyclopediaCristina DanaNo ratings yet