Download as pdf or txt
You might also like
- Bryophyta by O. P. SharmaDocument416 pagesBryophyta by O. P. Sharmaarba100% (19)
- Theory of Information ScienceDocument45 pagesTheory of Information ScienceriteshyuvarajNo ratings yet
- Combined Science PDFDocument57 pagesCombined Science PDFRadhika Meher58% (33)
- Philosophical FrameworksDocument9 pagesPhilosophical FrameworksmusharukwaNo ratings yet
- Quantum BiologicalDocument30 pagesQuantum BiologicalLe LeoNo ratings yet
- 4 13 2020 Scientists PDFDocument32 pages4 13 2020 Scientists PDFHeba NoiemNo ratings yet
- Disput Nitions: Ed DefiDocument6 pagesDisput Nitions: Ed DefiJohn Tyler KaufmanNo ratings yet
- On The Difference or Equality of Information MisinDocument15 pagesOn The Difference or Equality of Information Misingiulia.reis.vNo ratings yet
- Information Society NotesDocument16 pagesInformation Society NotesericNo ratings yet
- A Hombros de GigantesDocument38 pagesA Hombros de GigantesGaby PobletNo ratings yet
- A2010 What Is Science December. Pierre C HohenbergDocument8 pagesA2010 What Is Science December. Pierre C HohenbergCalvin Mona SandehangNo ratings yet
- Draper Conflict ThesisDocument6 pagesDraper Conflict Thesisheatherleeseattle100% (2)
- Deborah Avant The Ethics of Engaged Scholarship in ADocument22 pagesDeborah Avant The Ethics of Engaged Scholarship in AVespa SugioNo ratings yet
- Whatisinformation 03 00068 6Document25 pagesWhatisinformation 03 00068 6Demétrio Rocha PereiraNo ratings yet
- Evolution For Our Time: A Theory of Legal: Memetics 4Document42 pagesEvolution For Our Time: A Theory of Legal: Memetics 4Anahi VillegasNo ratings yet
- Weber Word 2001 BioScienceDocument9 pagesWeber Word 2001 BioSciencetagasalogNo ratings yet
- 1704 01614 PDFDocument8 pages1704 01614 PDFAdam ChabchoubNo ratings yet
- Topics in Contemporary EthicsDocument10 pagesTopics in Contemporary Ethicshutten7No ratings yet
- The Fourth Revolution Scientific Reductionism What Is A ParadigmDocument7 pagesThe Fourth Revolution Scientific Reductionism What Is A ParadigmMark Gutierrez100% (1)
- Case Chapter 3Document29 pagesCase Chapter 3globalspacestudiosNo ratings yet
- Neuroscience Education and The Evolution of The Human BrainDocument16 pagesNeuroscience Education and The Evolution of The Human BrainBale Muñoz CuartasNo ratings yet
- Epistemic Trust and The Ethics of Science Communication Against Transparency Openness Sincerity and HonestyDocument14 pagesEpistemic Trust and The Ethics of Science Communication Against Transparency Openness Sincerity and Honestyin doubtNo ratings yet
- Introduction: The Difference That Makes A Difference: David Chapman and Magnus RamageDocument5 pagesIntroduction: The Difference That Makes A Difference: David Chapman and Magnus RamageMickey ValleeNo ratings yet
- The University of Chicago Press Philosophy of Science AssociationDocument11 pagesThe University of Chicago Press Philosophy of Science AssociationAbuAbdur-RazzaqAl-MisriNo ratings yet
- The Social Psychology of "Pseudoscience": A Brief History: Arthur Still and Windy DrydenDocument26 pagesThe Social Psychology of "Pseudoscience": A Brief History: Arthur Still and Windy DrydenMalahat HajiyevaNo ratings yet
- Donald Maccrimmon MackayDocument5 pagesDonald Maccrimmon MackayDuan Gonmei100% (1)
- How Institutions Evolve: Evolutionary Theory and Institutional ChangeDocument27 pagesHow Institutions Evolve: Evolutionary Theory and Institutional ChangePam BarriosNo ratings yet
- Postmodern Possibility and The Convention On Biological DiversityDocument33 pagesPostmodern Possibility and The Convention On Biological DiversityweiNo ratings yet
- Mediated Modeling: A System Dynamics Approach To Environmental Consensus BuildingFrom EverandMediated Modeling: A System Dynamics Approach To Environmental Consensus BuildingNo ratings yet
- Bates-Fundamental Forms of InformationDocument13 pagesBates-Fundamental Forms of InformationthosampaNo ratings yet
- 5-Article Text-2683-1-10-20150416Document18 pages5-Article Text-2683-1-10-20150416PNo ratings yet
- Socsci Assay PDFDocument5 pagesSocsci Assay PDFRs Jyc RbdnNo ratings yet
- Ferguson Diversity in IR Theory Pluralism As An Opportunity For Understanding Global PoliticsDocument11 pagesFerguson Diversity in IR Theory Pluralism As An Opportunity For Understanding Global PoliticsRosarioNo ratings yet
- Extending The Extended Phenotype: Biology & Philosophy June 2004Document15 pagesExtending The Extended Phenotype: Biology & Philosophy June 2004AlejandroNo ratings yet
- Science Communication A Contemporary DefinitionDocument21 pagesScience Communication A Contemporary DefinitionLudmila NogueiraNo ratings yet
- TOK EssayDocument8 pagesTOK EssayRonakjain1998No ratings yet
- Module 1: Chapter 1: Introduction To Science Technology and Society SubtopicsDocument5 pagesModule 1: Chapter 1: Introduction To Science Technology and Society SubtopicsJonathan RebucasNo ratings yet
- Summer 2005 International Society For Environmental Ethics NewsletterDocument38 pagesSummer 2005 International Society For Environmental Ethics NewsletterInternational Society for Environmental EthicsNo ratings yet
- Information 2 DKFMDocument43 pagesInformation 2 DKFMglobalspacestudiosNo ratings yet
- In Defence of Generalized Darwinism - Aldrich, Hodgson, Hull, Knudsen, Mokyr, VanbergDocument20 pagesIn Defence of Generalized Darwinism - Aldrich, Hodgson, Hull, Knudsen, Mokyr, VanbergAndre VianaNo ratings yet
- Mark Burgin-Theory of Information (2009)Document688 pagesMark Burgin-Theory of Information (2009)losolores100% (2)
- Brust e Renck - 2021 - Judgment and Decision MakingDocument31 pagesBrust e Renck - 2021 - Judgment and Decision MakingsuwillowNo ratings yet
- Science and Pseudo-Science (Stanford Encyclopedia of Philosophy)Document15 pagesScience and Pseudo-Science (Stanford Encyclopedia of Philosophy)juanNo ratings yet
- Mooij, J. M. (2022) - Causality - From Data To ScienceDocument25 pagesMooij, J. M. (2022) - Causality - From Data To ScienceVictor Hugo Loaiza SeguraNo ratings yet
- Climbing Atop The Shoulders of Giants: The Impact of Institutions On Cumulative ResearchDocument32 pagesClimbing Atop The Shoulders of Giants: The Impact of Institutions On Cumulative ResearchBreno FragaNo ratings yet
- Lewenstein 2022Document4 pagesLewenstein 2022ludmilalqNo ratings yet
- Complexity Organization EvolutionandconstructallawDocument10 pagesComplexity Organization EvolutionandconstructallawGeorge PopescuNo ratings yet
- Research Paper On Long Term EvolutionDocument4 pagesResearch Paper On Long Term Evolutiongz7veyrh100% (1)
- Mat CHAPTER II 1 4 STSDocument16 pagesMat CHAPTER II 1 4 STSMarie DetarroNo ratings yet
- Chapter II - Science, Technology, and Society and The Human ConditionDocument16 pagesChapter II - Science, Technology, and Society and The Human ConditionJamesBuensalidoDellavaNo ratings yet
- Open Questions in Scientific Consensus Messaging ResearchDocument15 pagesOpen Questions in Scientific Consensus Messaging Researchin doubtNo ratings yet
- Science and PseudoscieneDocument19 pagesScience and PseudoscieneFriedrich AlexanderNo ratings yet
- Dominant Knowledge Systems and Local KnowledgeDocument25 pagesDominant Knowledge Systems and Local KnowledgeDepartment of Media Studies ST Thomas CollegeNo ratings yet
- Wilkins&Hull. Replication and ReproductionDocument17 pagesWilkins&Hull. Replication and ReproductionManu Rodríguez VázquezNo ratings yet
- Eng Ii-Eportfolio ReflectionDocument4 pagesEng Ii-Eportfolio Reflectionapi-611142808No ratings yet
- The Sciences of Science CommunicationDocument8 pagesThe Sciences of Science CommunicationHemantNo ratings yet
- Prescribed Title 6Document7 pagesPrescribed Title 6jszNo ratings yet
- The University of Chicago Press The History of Science SocietyDocument4 pagesThe University of Chicago Press The History of Science SocietyGertrudaNo ratings yet
- Life As Thermodynamic Evidence of Algorithmic Structure in Natural EnvironmentsDocument20 pagesLife As Thermodynamic Evidence of Algorithmic Structure in Natural EnvironmentsMohammed Samir KhokarNo ratings yet
- 1 s2.0 S187704281302908X Main PDFDocument5 pages1 s2.0 S187704281302908X Main PDFPuput Anistiya HarianiNo ratings yet
- Capurro Hjrland ConceptofinformationDocument71 pagesCapurro Hjrland ConceptofinformationlilianecrNo ratings yet
- Rose - 2012 - Democracy in The Contemporary Life Sciences-1Document14 pagesRose - 2012 - Democracy in The Contemporary Life Sciences-1Jaime Sebastián Cancino BarretoNo ratings yet
- Sex Education Final ExamDocument4 pagesSex Education Final Examapi-404291045No ratings yet
- Transformations Women Gender and Psychology 3rd Edition Crawford Test BankDocument14 pagesTransformations Women Gender and Psychology 3rd Edition Crawford Test Bankmatthewelmerwjxqf100% (22)
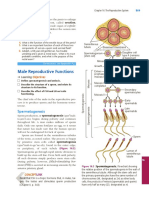
- Male Reproductive Functions: Did You Get It?Document1 pageMale Reproductive Functions: Did You Get It?Krizziane Ivy CuajotorNo ratings yet
- Sex and Context - Hormones and Primate Sexual MotivationDocument19 pagesSex and Context - Hormones and Primate Sexual MotivationbobbyNo ratings yet
- Sex and Love 2Document11 pagesSex and Love 2kojo2kgNo ratings yet
- Coordinated Functions of The Nervous, Endocrine, and Reproductive SystemsDocument30 pagesCoordinated Functions of The Nervous, Endocrine, and Reproductive SystemsSarah CruzNo ratings yet
- Responsible Parenthood: Role of Father & Mother in The FamilyDocument7 pagesResponsible Parenthood: Role of Father & Mother in The FamilyBhenjo Hernandez BaronaNo ratings yet
- Premarital SexDocument4 pagesPremarital Sexpedring_ganadenNo ratings yet
- CP102AREPORTING-Natural Family Planning (NFP) MethodsDocument31 pagesCP102AREPORTING-Natural Family Planning (NFP) MethodsCHONANo ratings yet
- The Sympto-Thermal Method - Ten Years of Change PDFDocument18 pagesThe Sympto-Thermal Method - Ten Years of Change PDFFabian GuardaNo ratings yet
- Kuliah 3. Porifera PDFDocument29 pagesKuliah 3. Porifera PDFSamuel Ivana NaibahoNo ratings yet
- Types of Family Planning MethodsDocument6 pagesTypes of Family Planning MethodsJenny Soriano100% (1)
- What Is The Creighton Model?Document2 pagesWhat Is The Creighton Model?Amy ZulliantiNo ratings yet
- Menstrual CycleDocument6 pagesMenstrual CycleKyndra Maxynne CamachoNo ratings yet
- Erogenous Zone: 1 ClassificationDocument6 pagesErogenous Zone: 1 Classificationnathan100% (2)
- Artificial Insemination - Docx1Document3 pagesArtificial Insemination - Docx1carol ann saliseNo ratings yet
- B1.3.5 Menstrual Cycle TimelineDocument1 pageB1.3.5 Menstrual Cycle TimelineDearbhla HubbardNo ratings yet
- SDKI 2002 - Kuesioner PriaDocument19 pagesSDKI 2002 - Kuesioner Priakiki rizkiNo ratings yet
- Final Draft Research PaperDocument10 pagesFinal Draft Research Paperapi-508926158No ratings yet
- The Problem of Defining The FamilyDocument27 pagesThe Problem of Defining The FamilyJuvy BasilioNo ratings yet
- 2021 Report: Scientific, Legal, Pro-Life, & Pro-Choice Sources On When A Human's Life BeginsDocument209 pages2021 Report: Scientific, Legal, Pro-Life, & Pro-Choice Sources On When A Human's Life BeginsSteve JacobsNo ratings yet
- Sex Gender and SexualityDocument15 pagesSex Gender and SexualityGeorge Aryan Dela VegaNo ratings yet
- Unpacking The Self - Sexual Self: Prepared By: Marisol ArizaDocument29 pagesUnpacking The Self - Sexual Self: Prepared By: Marisol ArizaJohn Kenneth ConcepcionNo ratings yet
- Using Musculoskeletal Models To Explore Strategies For Improving Performance in Electrical Stimulation-Induced Leg Cycle ErgometryDocument196 pagesUsing Musculoskeletal Models To Explore Strategies For Improving Performance in Electrical Stimulation-Induced Leg Cycle ErgometryAna de SousaNo ratings yet
- Estrous Cycle Signs of Estrous Artificial Insemination Merits and Demerits of AI Embryo Transfer TechnologyDocument88 pagesEstrous Cycle Signs of Estrous Artificial Insemination Merits and Demerits of AI Embryo Transfer Technologyshubh100% (1)
- Sexual SelfDocument54 pagesSexual SelfCidrick CorpuzNo ratings yet
- 46 Animal ReproductionDocument18 pages46 Animal ReproductionДина МедетбековаNo ratings yet
- Labadia, John Daniel ScienceDocument2 pagesLabadia, John Daniel ScienceJohn Daniel LabadiaNo ratings yet