
Mercury in The Diatoms of Various Ecological Forma
Mercury in The Diatoms of Various Ecological Forma
You might also like
- Pre-Commissioning and CommissioningDocument9 pagesPre-Commissioning and CommissioningSaurabh Sharma100% (7)
- The First Ray by The Ascended Master El Morya (1954)Document121 pagesThe First Ray by The Ascended Master El Morya (1954)John LeeNo ratings yet
- 1 s2.0 S0025326X22008116 MainDocument10 pages1 s2.0 S0025326X22008116 MainyosyNo ratings yet
- 1 s2.0 S0025326X22000339 MainDocument10 pages1 s2.0 S0025326X22000339 MainOscar Herrera AmézquitaNo ratings yet
- Shipborne Nutrient Dynamics and Impact On The Eutrophication in The Baltic Sea 2019Document19 pagesShipborne Nutrient Dynamics and Impact On The Eutrophication in The Baltic Sea 2019Paola VillanuevaNo ratings yet
- 2021boletn de Contaminacin MarinaDocument12 pages2021boletn de Contaminacin Marinarafael mezaNo ratings yet
- Co Occurrence of Contaminants in Marine Fish From The Nort - 2021 - EnvironmentDocument13 pagesCo Occurrence of Contaminants in Marine Fish From The Nort - 2021 - EnvironmentAlexander VassilevNo ratings yet
- 1 PBDocument8 pages1 PBElena NicolauNo ratings yet
- Limnologica: Wojciech Ejankowski, Tomasz LenardDocument8 pagesLimnologica: Wojciech Ejankowski, Tomasz LenardMoltimer Folchart CrawNo ratings yet
- Chapter One 1.0 1.1 Polychlorinated Biphenyls (PCBS)Document10 pagesChapter One 1.0 1.1 Polychlorinated Biphenyls (PCBS)Akpan EkomNo ratings yet
- HG Mining Ponds Colombia-MainDocument13 pagesHG Mining Ponds Colombia-Maingrecia.valdiviaNo ratings yet
- Bourgeois Et Al 2017 GBCDocument24 pagesBourgeois Et Al 2017 GBCSixto Gutiérrez SaavedraNo ratings yet
- García-Pimentel 2023-1Document17 pagesGarcía-Pimentel 2023-1Eduardo Chavez MontesNo ratings yet
- Lazar Et Al - The Influence of Climate Changes On Heavy Metals Accumulation in Alosa ImmaculataDocument14 pagesLazar Et Al - The Influence of Climate Changes On Heavy Metals Accumulation in Alosa ImmaculataNinaNicoletaNo ratings yet
- AinaDocument16 pagesAinaRodrigue AdechinaNo ratings yet
- Zooplankton From A North Western Mediterranean Area As A Model of Metal Transfer in A Marine EnvironmentDocument12 pagesZooplankton From A North Western Mediterranean Area As A Model of Metal Transfer in A Marine EnvironmentFloraNo ratings yet
- Ecological EngineeringDocument12 pagesEcological EngineeringRahmat PangestuNo ratings yet
- Bojarczuk2018 Article TheEffectOfAnthropogenicAndNatDocument13 pagesBojarczuk2018 Article TheEffectOfAnthropogenicAndNatelias bello leonNo ratings yet
- JGR Biogeosciences - 2019 - Vergara Jara - Seasonal Changes in Carbonate Saturation State and Air Sea CO2 Fluxes During AnDocument15 pagesJGR Biogeosciences - 2019 - Vergara Jara - Seasonal Changes in Carbonate Saturation State and Air Sea CO2 Fluxes During Anmarco antonio pinto torresNo ratings yet
- Tsyban 1999Document6 pagesTsyban 1999Rahul AmbawataNo ratings yet
- Fmars 09 864306Document11 pagesFmars 09 864306Carolina SotomayorNo ratings yet
- Behaviors of Nitrogen, Iron and Sulfur Compounds in Contaminated Marine SedimentDocument7 pagesBehaviors of Nitrogen, Iron and Sulfur Compounds in Contaminated Marine SedimentZahed AnsaryNo ratings yet
- Contaminantes Orgánicos en Las Aguas ProfundasDocument13 pagesContaminantes Orgánicos en Las Aguas Profundasraquel mr18No ratings yet
- Photosynthesis and Respiration of Baltic Sea BenthDocument19 pagesPhotosynthesis and Respiration of Baltic Sea BenthAyan DuttaNo ratings yet
- Moreno-González - 2013 - Seasonal Input of Regulated and Emerging Organic Pollutants Through-1Document1 pageMoreno-González - 2013 - Seasonal Input of Regulated and Emerging Organic Pollutants Through-1Rubén Moreno GonzálezNo ratings yet
- OCR Suppl 2022-0075Document35 pagesOCR Suppl 2022-0075luissecundesNo ratings yet
- Ocean Fertilization: A Scientific Summary For Policy MakersDocument20 pagesOcean Fertilization: A Scientific Summary For Policy MakersDaniel Sanchez DiazNo ratings yet
- Aguas de Mina SalinasDocument11 pagesAguas de Mina SalinasBernardo Gamarra ValdiviezoNo ratings yet
- Aydin-Onen Ve Diğ., 2012Document9 pagesAydin-Onen Ve Diğ., 2012Sinem AydınNo ratings yet
- FSZ 219Document12 pagesFSZ 219Ebenezer PasaribuNo ratings yet
- Impact of Anthropogenic Activity and Lusi-Mud VolcDocument7 pagesImpact of Anthropogenic Activity and Lusi-Mud VolcShashanth VenkatesanNo ratings yet
- Assessment of Heavy Metal Contamination of River Gora Kaduna, NigeriaDocument5 pagesAssessment of Heavy Metal Contamination of River Gora Kaduna, NigeriaToheeb AtandaNo ratings yet
- Science of The Total Environment: Contents Lists Available atDocument12 pagesScience of The Total Environment: Contents Lists Available atLucas B. TheodoroviezNo ratings yet
- Science of The Total Environment: Contents Lists Available atDocument12 pagesScience of The Total Environment: Contents Lists Available atDavi BarbosaNo ratings yet
- Dissolved Organic Matter DOM Release by Phytoplankton in The Contemporary and Future OceanDocument28 pagesDissolved Organic Matter DOM Release by Phytoplankton in The Contemporary and Future OceanFabio RicciNo ratings yet
- Stable Isotope Analysis of Food Web Structure and PDFDocument17 pagesStable Isotope Analysis of Food Web Structure and PDFAna Jimena Celis MartínezNo ratings yet
- Envi Toxicity in Baltic SeaDocument8 pagesEnvi Toxicity in Baltic SeaAleine Leilanie OroNo ratings yet
- COPs en BallenasDocument8 pagesCOPs en BallenasFrancisco JavierNo ratings yet
- Lake Como Contaminated With Chemicals Banned in The 1970sDocument2 pagesLake Como Contaminated With Chemicals Banned in The 1970sDemoNo ratings yet
- Matunguru - Okito Et Al 2022 - Assessment of Physicochemial Parametrs in Relation With Ecology of Bagrus in Lake Albert-DRCDocument18 pagesMatunguru - Okito Et Al 2022 - Assessment of Physicochemial Parametrs in Relation With Ecology of Bagrus in Lake Albert-DRCGabriel MUKABO OKITONo ratings yet
- High CO2 and Warming Affect Microzooplankton FoodDocument18 pagesHigh CO2 and Warming Affect Microzooplankton Foodadityakapuriya11No ratings yet
- Assessment of Selected Heavy Metals (CD, CR, Cu, Ni and PB) Concentration inDocument8 pagesAssessment of Selected Heavy Metals (CD, CR, Cu, Ni and PB) Concentration inInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Influence of Salinity and Rare Earth Elements On Simultaneous Removal of CD, CR, Cu, HG, Ni and PB From Contaminated Waters by Living MacroalgaeDocument13 pagesInfluence of Salinity and Rare Earth Elements On Simultaneous Removal of CD, CR, Cu, HG, Ni and PB From Contaminated Waters by Living MacroalgaeBrigita de BrillarNo ratings yet
- Journal of Experimental Marine Biology and Ecology: SciencedirectDocument12 pagesJournal of Experimental Marine Biology and Ecology: SciencedirectTalitha RdmNo ratings yet
- Marine Pollution Bulletin Volume 95 Issue 1 2015 (Doi 10.1016 - J.marpolbul.2015.03.035) Zhang, Ling Shi, Zhen Zhang, JingPing Jiang, Zhijian Wang, F - Spatial and Seasonal Characteristics of DisDocument8 pagesMarine Pollution Bulletin Volume 95 Issue 1 2015 (Doi 10.1016 - J.marpolbul.2015.03.035) Zhang, Ling Shi, Zhen Zhang, JingPing Jiang, Zhijian Wang, F - Spatial and Seasonal Characteristics of DisJoao Finisterre GomezNo ratings yet
- Baseline Evaluation of Metal Contamination in Teleost Fishes - 2019 - Marine PolDocument8 pagesBaseline Evaluation of Metal Contamination in Teleost Fishes - 2019 - Marine Polabdelmonim.haidourNo ratings yet
- Ocean Acidification: A Hidden Risk For Sustainable DevelopmentDocument4 pagesOcean Acidification: A Hidden Risk For Sustainable DevelopmentSir TemplarNo ratings yet
- 1 s2.0 S0078323417300726 MainDocument13 pages1 s2.0 S0078323417300726 MainRahmaNo ratings yet
- Aslam 2016Document19 pagesAslam 2016Larissa Risky AmaliaNo ratings yet
- HHJDocument13 pagesHHJԳոռ ԽաչատրյանNo ratings yet
- Basin Specific Changes in Filamentous Cyanobacteria Community C - 2020 - HarmfulDocument12 pagesBasin Specific Changes in Filamentous Cyanobacteria Community C - 2020 - Harmfulzan zibarNo ratings yet
- 1 s2.0 S0048969722057825 MainDocument6 pages1 s2.0 S0048969722057825 MainMuhammad FadliNo ratings yet
- Bloomalgas Verano 2016Document10 pagesBloomalgas Verano 2016Cesar DuranNo ratings yet
- 2023 Article 26317Document18 pages2023 Article 26317mariaNo ratings yet
- Biola 2017 PDFDocument12 pagesBiola 2017 PDFBob AnggaraNo ratings yet
- 1 s2.0 S0048969719343839 MainDocument17 pages1 s2.0 S0048969719343839 MainArvind NegiNo ratings yet
- FNX 214Document12 pagesFNX 214Muhammad FikriNo ratings yet
- Mitchelmore Et Al., (2019)Document13 pagesMitchelmore Et Al., (2019)Asistente LaboratorioNo ratings yet
- Journal Pre-Proof: Science of The Total EnvironmentDocument42 pagesJournal Pre-Proof: Science of The Total Environmentmaristela vargasNo ratings yet
- 1-S2.0-S2352485516302390-Main SKLMP Proposal Jan23Document7 pages1-S2.0-S2352485516302390-Main SKLMP Proposal Jan23Martin TsuiNo ratings yet
- The Tragedy of the Commodity: Oceans, Fisheries, and AquacultureFrom EverandThe Tragedy of the Commodity: Oceans, Fisheries, and AquacultureNo ratings yet
- Blue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and ImplementationFrom EverandBlue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and ImplementationTomohiro KuwaeNo ratings yet
- DiatomsDocument10 pagesDiatomsCatalina SarriaNo ratings yet
- BioremediacionDocument8 pagesBioremediacionCatalina SarriaNo ratings yet
- Absorbancia PDFDocument1 pageAbsorbancia PDFCatalina SarriaNo ratings yet
- Reflectancia PDFDocument1 pageReflectancia PDFCatalina SarriaNo ratings yet
- Content Mapping Template - HubSpotDocument7 pagesContent Mapping Template - HubSpotCatalina SarriaNo ratings yet
- Estimacion de CostosDocument44 pagesEstimacion de CostosCatalina SarriaNo ratings yet
- Plant LifeDocument19 pagesPlant Lifesheena2sa100% (3)
- Ujian Sains Tingkatan 2Document15 pagesUjian Sains Tingkatan 2SITI ZAIDAH AHMAD75% (12)
- Walko Et Al. - 2000 - Coupled Atmosphere-Biophysics-Hydrology Models For Environmental ModelingDocument14 pagesWalko Et Al. - 2000 - Coupled Atmosphere-Biophysics-Hydrology Models For Environmental ModelingRocio Veronica Gutierrez RosalesNo ratings yet
- De Thi Giua Ki 2 Tieng Anh 8 Global SuccessDocument9 pagesDe Thi Giua Ki 2 Tieng Anh 8 Global SuccessQuỳnh PhạmNo ratings yet
- Photosynthesis Introduction WorksheetDocument3 pagesPhotosynthesis Introduction WorksheetGaboOrtizVieiraNo ratings yet
- Reconstruction of The Evolution of The Alpine-Himalayan OrogenDocument3 pagesReconstruction of The Evolution of The Alpine-Himalayan Orogentamir chongNo ratings yet
- ABEn 145 - Tribunalo - Lab 6Document10 pagesABEn 145 - Tribunalo - Lab 6HeliNo ratings yet
- Force and PressureDocument6 pagesForce and PressureRaj KumarNo ratings yet
- Power System Engineering Lecture 1Document16 pagesPower System Engineering Lecture 1nnvpratapNo ratings yet
- BiodiversityDocument3 pagesBiodiversityJóhann G. ThorarensenNo ratings yet
- Chemical Engineering ObjectiveDocument3 pagesChemical Engineering ObjectiverastamanrmNo ratings yet
- Effects of Boron Doping in Low - and High-surface-Area Carbon PowdersDocument12 pagesEffects of Boron Doping in Low - and High-surface-Area Carbon Powdersvictor romeroNo ratings yet
- PIPE 180 Updated 2015Document180 pagesPIPE 180 Updated 2015joe1256No ratings yet
- Tutorial 1Document3 pagesTutorial 1doldolzzNo ratings yet
- Green Engine Report FinalDocument12 pagesGreen Engine Report FinalPurneshwari Devi100% (2)
- Design and Simulation of Maximum Power Point Tracking of Photovoltaic System Using ANNDocument5 pagesDesign and Simulation of Maximum Power Point Tracking of Photovoltaic System Using ANNMd Samiul Haque SunnyNo ratings yet
- X-Ray Transmission and Moseley's LawDocument6 pagesX-Ray Transmission and Moseley's LawFaiz ZinNo ratings yet
- Class 7 - Air Around UsDocument3 pagesClass 7 - Air Around Usmamta sehrawat100% (1)
- ReadingDocument3 pagesReadingОля БондаренкоNo ratings yet
- Hydrosphere and BiosphereDocument46 pagesHydrosphere and BiosphereZelie GuerinNo ratings yet
- NSTPDocument6 pagesNSTPRyzajane DalingNo ratings yet
- Lesson Objectives: Knows The Different Types of Containment Covered by The Term "Packaged Form"Document41 pagesLesson Objectives: Knows The Different Types of Containment Covered by The Term "Packaged Form"Farid Muhamad100% (6)
- Lecture 5 HvacDocument42 pagesLecture 5 HvacJohn Bennett100% (2)
- Biochar-Based Nano-Composites For The Decontamination of Wastewater A ReviewDocument16 pagesBiochar-Based Nano-Composites For The Decontamination of Wastewater A ReviewNa ZhangNo ratings yet
- Concept Note 1Document4 pagesConcept Note 1Sonali SharmaNo ratings yet
- Icse Specimen QP - Environmental ScienceDocument9 pagesIcse Specimen QP - Environmental ScienceLOKESH KUMARNo ratings yet
- Huang Et Al. - 2019 - Modeling and Energy Management of A Photovoltaic Fuel Cell Battery HybridDocument14 pagesHuang Et Al. - 2019 - Modeling and Energy Management of A Photovoltaic Fuel Cell Battery HybridJAVARIA ABDUR REHMANNo ratings yet
- Topic 5 TestDocument4 pagesTopic 5 TestGenevieve LawrenceNo ratings yet
































































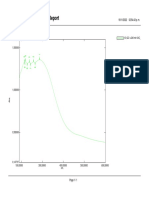
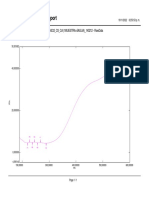






























Download as pdf or txt
You might also like
- Pre-Commissioning and CommissioningDocument9 pagesPre-Commissioning and CommissioningSaurabh Sharma100% (7)
- The First Ray by The Ascended Master El Morya (1954)Document121 pagesThe First Ray by The Ascended Master El Morya (1954)John LeeNo ratings yet
- 1 s2.0 S0025326X22008116 MainDocument10 pages1 s2.0 S0025326X22008116 MainyosyNo ratings yet
- 1 s2.0 S0025326X22000339 MainDocument10 pages1 s2.0 S0025326X22000339 MainOscar Herrera AmézquitaNo ratings yet
- Shipborne Nutrient Dynamics and Impact On The Eutrophication in The Baltic Sea 2019Document19 pagesShipborne Nutrient Dynamics and Impact On The Eutrophication in The Baltic Sea 2019Paola VillanuevaNo ratings yet
- 2021boletn de Contaminacin MarinaDocument12 pages2021boletn de Contaminacin Marinarafael mezaNo ratings yet
- Co Occurrence of Contaminants in Marine Fish From The Nort - 2021 - EnvironmentDocument13 pagesCo Occurrence of Contaminants in Marine Fish From The Nort - 2021 - EnvironmentAlexander VassilevNo ratings yet
- 1 PBDocument8 pages1 PBElena NicolauNo ratings yet
- Limnologica: Wojciech Ejankowski, Tomasz LenardDocument8 pagesLimnologica: Wojciech Ejankowski, Tomasz LenardMoltimer Folchart CrawNo ratings yet
- Chapter One 1.0 1.1 Polychlorinated Biphenyls (PCBS)Document10 pagesChapter One 1.0 1.1 Polychlorinated Biphenyls (PCBS)Akpan EkomNo ratings yet
- HG Mining Ponds Colombia-MainDocument13 pagesHG Mining Ponds Colombia-Maingrecia.valdiviaNo ratings yet
- Bourgeois Et Al 2017 GBCDocument24 pagesBourgeois Et Al 2017 GBCSixto Gutiérrez SaavedraNo ratings yet
- García-Pimentel 2023-1Document17 pagesGarcía-Pimentel 2023-1Eduardo Chavez MontesNo ratings yet
- Lazar Et Al - The Influence of Climate Changes On Heavy Metals Accumulation in Alosa ImmaculataDocument14 pagesLazar Et Al - The Influence of Climate Changes On Heavy Metals Accumulation in Alosa ImmaculataNinaNicoletaNo ratings yet
- AinaDocument16 pagesAinaRodrigue AdechinaNo ratings yet
- Zooplankton From A North Western Mediterranean Area As A Model of Metal Transfer in A Marine EnvironmentDocument12 pagesZooplankton From A North Western Mediterranean Area As A Model of Metal Transfer in A Marine EnvironmentFloraNo ratings yet
- Ecological EngineeringDocument12 pagesEcological EngineeringRahmat PangestuNo ratings yet
- Bojarczuk2018 Article TheEffectOfAnthropogenicAndNatDocument13 pagesBojarczuk2018 Article TheEffectOfAnthropogenicAndNatelias bello leonNo ratings yet
- JGR Biogeosciences - 2019 - Vergara Jara - Seasonal Changes in Carbonate Saturation State and Air Sea CO2 Fluxes During AnDocument15 pagesJGR Biogeosciences - 2019 - Vergara Jara - Seasonal Changes in Carbonate Saturation State and Air Sea CO2 Fluxes During Anmarco antonio pinto torresNo ratings yet
- Tsyban 1999Document6 pagesTsyban 1999Rahul AmbawataNo ratings yet
- Fmars 09 864306Document11 pagesFmars 09 864306Carolina SotomayorNo ratings yet
- Behaviors of Nitrogen, Iron and Sulfur Compounds in Contaminated Marine SedimentDocument7 pagesBehaviors of Nitrogen, Iron and Sulfur Compounds in Contaminated Marine SedimentZahed AnsaryNo ratings yet
- Contaminantes Orgánicos en Las Aguas ProfundasDocument13 pagesContaminantes Orgánicos en Las Aguas Profundasraquel mr18No ratings yet
- Photosynthesis and Respiration of Baltic Sea BenthDocument19 pagesPhotosynthesis and Respiration of Baltic Sea BenthAyan DuttaNo ratings yet
- Moreno-González - 2013 - Seasonal Input of Regulated and Emerging Organic Pollutants Through-1Document1 pageMoreno-González - 2013 - Seasonal Input of Regulated and Emerging Organic Pollutants Through-1Rubén Moreno GonzálezNo ratings yet
- OCR Suppl 2022-0075Document35 pagesOCR Suppl 2022-0075luissecundesNo ratings yet
- Ocean Fertilization: A Scientific Summary For Policy MakersDocument20 pagesOcean Fertilization: A Scientific Summary For Policy MakersDaniel Sanchez DiazNo ratings yet
- Aguas de Mina SalinasDocument11 pagesAguas de Mina SalinasBernardo Gamarra ValdiviezoNo ratings yet
- Aydin-Onen Ve Diğ., 2012Document9 pagesAydin-Onen Ve Diğ., 2012Sinem AydınNo ratings yet
- FSZ 219Document12 pagesFSZ 219Ebenezer PasaribuNo ratings yet
- Impact of Anthropogenic Activity and Lusi-Mud VolcDocument7 pagesImpact of Anthropogenic Activity and Lusi-Mud VolcShashanth VenkatesanNo ratings yet
- Assessment of Heavy Metal Contamination of River Gora Kaduna, NigeriaDocument5 pagesAssessment of Heavy Metal Contamination of River Gora Kaduna, NigeriaToheeb AtandaNo ratings yet
- Science of The Total Environment: Contents Lists Available atDocument12 pagesScience of The Total Environment: Contents Lists Available atLucas B. TheodoroviezNo ratings yet
- Science of The Total Environment: Contents Lists Available atDocument12 pagesScience of The Total Environment: Contents Lists Available atDavi BarbosaNo ratings yet
- Dissolved Organic Matter DOM Release by Phytoplankton in The Contemporary and Future OceanDocument28 pagesDissolved Organic Matter DOM Release by Phytoplankton in The Contemporary and Future OceanFabio RicciNo ratings yet
- Stable Isotope Analysis of Food Web Structure and PDFDocument17 pagesStable Isotope Analysis of Food Web Structure and PDFAna Jimena Celis MartínezNo ratings yet
- Envi Toxicity in Baltic SeaDocument8 pagesEnvi Toxicity in Baltic SeaAleine Leilanie OroNo ratings yet
- COPs en BallenasDocument8 pagesCOPs en BallenasFrancisco JavierNo ratings yet
- Lake Como Contaminated With Chemicals Banned in The 1970sDocument2 pagesLake Como Contaminated With Chemicals Banned in The 1970sDemoNo ratings yet
- Matunguru - Okito Et Al 2022 - Assessment of Physicochemial Parametrs in Relation With Ecology of Bagrus in Lake Albert-DRCDocument18 pagesMatunguru - Okito Et Al 2022 - Assessment of Physicochemial Parametrs in Relation With Ecology of Bagrus in Lake Albert-DRCGabriel MUKABO OKITONo ratings yet
- High CO2 and Warming Affect Microzooplankton FoodDocument18 pagesHigh CO2 and Warming Affect Microzooplankton Foodadityakapuriya11No ratings yet
- Assessment of Selected Heavy Metals (CD, CR, Cu, Ni and PB) Concentration inDocument8 pagesAssessment of Selected Heavy Metals (CD, CR, Cu, Ni and PB) Concentration inInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Influence of Salinity and Rare Earth Elements On Simultaneous Removal of CD, CR, Cu, HG, Ni and PB From Contaminated Waters by Living MacroalgaeDocument13 pagesInfluence of Salinity and Rare Earth Elements On Simultaneous Removal of CD, CR, Cu, HG, Ni and PB From Contaminated Waters by Living MacroalgaeBrigita de BrillarNo ratings yet
- Journal of Experimental Marine Biology and Ecology: SciencedirectDocument12 pagesJournal of Experimental Marine Biology and Ecology: SciencedirectTalitha RdmNo ratings yet
- Marine Pollution Bulletin Volume 95 Issue 1 2015 (Doi 10.1016 - J.marpolbul.2015.03.035) Zhang, Ling Shi, Zhen Zhang, JingPing Jiang, Zhijian Wang, F - Spatial and Seasonal Characteristics of DisDocument8 pagesMarine Pollution Bulletin Volume 95 Issue 1 2015 (Doi 10.1016 - J.marpolbul.2015.03.035) Zhang, Ling Shi, Zhen Zhang, JingPing Jiang, Zhijian Wang, F - Spatial and Seasonal Characteristics of DisJoao Finisterre GomezNo ratings yet
- Baseline Evaluation of Metal Contamination in Teleost Fishes - 2019 - Marine PolDocument8 pagesBaseline Evaluation of Metal Contamination in Teleost Fishes - 2019 - Marine Polabdelmonim.haidourNo ratings yet
- Ocean Acidification: A Hidden Risk For Sustainable DevelopmentDocument4 pagesOcean Acidification: A Hidden Risk For Sustainable DevelopmentSir TemplarNo ratings yet
- 1 s2.0 S0078323417300726 MainDocument13 pages1 s2.0 S0078323417300726 MainRahmaNo ratings yet
- Aslam 2016Document19 pagesAslam 2016Larissa Risky AmaliaNo ratings yet
- HHJDocument13 pagesHHJԳոռ ԽաչատրյանNo ratings yet
- Basin Specific Changes in Filamentous Cyanobacteria Community C - 2020 - HarmfulDocument12 pagesBasin Specific Changes in Filamentous Cyanobacteria Community C - 2020 - Harmfulzan zibarNo ratings yet
- 1 s2.0 S0048969722057825 MainDocument6 pages1 s2.0 S0048969722057825 MainMuhammad FadliNo ratings yet
- Bloomalgas Verano 2016Document10 pagesBloomalgas Verano 2016Cesar DuranNo ratings yet
- 2023 Article 26317Document18 pages2023 Article 26317mariaNo ratings yet
- Biola 2017 PDFDocument12 pagesBiola 2017 PDFBob AnggaraNo ratings yet
- 1 s2.0 S0048969719343839 MainDocument17 pages1 s2.0 S0048969719343839 MainArvind NegiNo ratings yet
- FNX 214Document12 pagesFNX 214Muhammad FikriNo ratings yet
- Mitchelmore Et Al., (2019)Document13 pagesMitchelmore Et Al., (2019)Asistente LaboratorioNo ratings yet
- Journal Pre-Proof: Science of The Total EnvironmentDocument42 pagesJournal Pre-Proof: Science of The Total Environmentmaristela vargasNo ratings yet
- 1-S2.0-S2352485516302390-Main SKLMP Proposal Jan23Document7 pages1-S2.0-S2352485516302390-Main SKLMP Proposal Jan23Martin TsuiNo ratings yet
- The Tragedy of the Commodity: Oceans, Fisheries, and AquacultureFrom EverandThe Tragedy of the Commodity: Oceans, Fisheries, and AquacultureNo ratings yet
- Blue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and ImplementationFrom EverandBlue Carbon in Shallow Coastal Ecosystems: Carbon Dynamics, Policy, and ImplementationTomohiro KuwaeNo ratings yet
- DiatomsDocument10 pagesDiatomsCatalina SarriaNo ratings yet
- BioremediacionDocument8 pagesBioremediacionCatalina SarriaNo ratings yet
- Absorbancia PDFDocument1 pageAbsorbancia PDFCatalina SarriaNo ratings yet
- Reflectancia PDFDocument1 pageReflectancia PDFCatalina SarriaNo ratings yet
- Content Mapping Template - HubSpotDocument7 pagesContent Mapping Template - HubSpotCatalina SarriaNo ratings yet
- Estimacion de CostosDocument44 pagesEstimacion de CostosCatalina SarriaNo ratings yet
- Plant LifeDocument19 pagesPlant Lifesheena2sa100% (3)
- Ujian Sains Tingkatan 2Document15 pagesUjian Sains Tingkatan 2SITI ZAIDAH AHMAD75% (12)
- Walko Et Al. - 2000 - Coupled Atmosphere-Biophysics-Hydrology Models For Environmental ModelingDocument14 pagesWalko Et Al. - 2000 - Coupled Atmosphere-Biophysics-Hydrology Models For Environmental ModelingRocio Veronica Gutierrez RosalesNo ratings yet
- De Thi Giua Ki 2 Tieng Anh 8 Global SuccessDocument9 pagesDe Thi Giua Ki 2 Tieng Anh 8 Global SuccessQuỳnh PhạmNo ratings yet
- Photosynthesis Introduction WorksheetDocument3 pagesPhotosynthesis Introduction WorksheetGaboOrtizVieiraNo ratings yet
- Reconstruction of The Evolution of The Alpine-Himalayan OrogenDocument3 pagesReconstruction of The Evolution of The Alpine-Himalayan Orogentamir chongNo ratings yet
- ABEn 145 - Tribunalo - Lab 6Document10 pagesABEn 145 - Tribunalo - Lab 6HeliNo ratings yet
- Force and PressureDocument6 pagesForce and PressureRaj KumarNo ratings yet
- Power System Engineering Lecture 1Document16 pagesPower System Engineering Lecture 1nnvpratapNo ratings yet
- BiodiversityDocument3 pagesBiodiversityJóhann G. ThorarensenNo ratings yet
- Chemical Engineering ObjectiveDocument3 pagesChemical Engineering ObjectiverastamanrmNo ratings yet
- Effects of Boron Doping in Low - and High-surface-Area Carbon PowdersDocument12 pagesEffects of Boron Doping in Low - and High-surface-Area Carbon Powdersvictor romeroNo ratings yet
- PIPE 180 Updated 2015Document180 pagesPIPE 180 Updated 2015joe1256No ratings yet
- Tutorial 1Document3 pagesTutorial 1doldolzzNo ratings yet
- Green Engine Report FinalDocument12 pagesGreen Engine Report FinalPurneshwari Devi100% (2)
- Design and Simulation of Maximum Power Point Tracking of Photovoltaic System Using ANNDocument5 pagesDesign and Simulation of Maximum Power Point Tracking of Photovoltaic System Using ANNMd Samiul Haque SunnyNo ratings yet
- X-Ray Transmission and Moseley's LawDocument6 pagesX-Ray Transmission and Moseley's LawFaiz ZinNo ratings yet
- Class 7 - Air Around UsDocument3 pagesClass 7 - Air Around Usmamta sehrawat100% (1)
- ReadingDocument3 pagesReadingОля БондаренкоNo ratings yet
- Hydrosphere and BiosphereDocument46 pagesHydrosphere and BiosphereZelie GuerinNo ratings yet
- NSTPDocument6 pagesNSTPRyzajane DalingNo ratings yet
- Lesson Objectives: Knows The Different Types of Containment Covered by The Term "Packaged Form"Document41 pagesLesson Objectives: Knows The Different Types of Containment Covered by The Term "Packaged Form"Farid Muhamad100% (6)
- Lecture 5 HvacDocument42 pagesLecture 5 HvacJohn Bennett100% (2)
- Biochar-Based Nano-Composites For The Decontamination of Wastewater A ReviewDocument16 pagesBiochar-Based Nano-Composites For The Decontamination of Wastewater A ReviewNa ZhangNo ratings yet
- Concept Note 1Document4 pagesConcept Note 1Sonali SharmaNo ratings yet
- Icse Specimen QP - Environmental ScienceDocument9 pagesIcse Specimen QP - Environmental ScienceLOKESH KUMARNo ratings yet
- Huang Et Al. - 2019 - Modeling and Energy Management of A Photovoltaic Fuel Cell Battery HybridDocument14 pagesHuang Et Al. - 2019 - Modeling and Energy Management of A Photovoltaic Fuel Cell Battery HybridJAVARIA ABDUR REHMANNo ratings yet
- Topic 5 TestDocument4 pagesTopic 5 TestGenevieve LawrenceNo ratings yet