
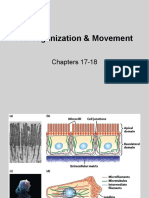
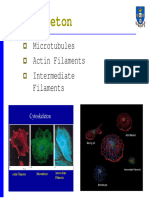
The Cytoskeleton
The Cytoskeleton
You might also like
- University of Texas BIO 325 ExamDocument6 pagesUniversity of Texas BIO 325 ExamGeetika InstaNo ratings yet
- Kami Export - Worksheet 14 Central DogmaDocument2 pagesKami Export - Worksheet 14 Central DogmaLindsey TamlinNo ratings yet
- 11.4 Meiosis: Multiple ChoiceDocument5 pages11.4 Meiosis: Multiple ChoiceMING ZHUNo ratings yet
- Amino Acid MetabolsmDocument16 pagesAmino Acid MetabolsmManila MedNo ratings yet
- 17&18) Cell Organization & MovementDocument28 pages17&18) Cell Organization & MovementRNo ratings yet
- BY2201 EmmaC CytoskeletonIandIIDocument62 pagesBY2201 EmmaC CytoskeletonIandIIRebecca ReillyNo ratings yet
- The CytoskeletonDocument10 pagesThe CytoskeletonAhmed khanNo ratings yet
- 2 Cytoskeleton & Cell MotilityDocument47 pages2 Cytoskeleton & Cell MotilityChristineNo ratings yet
- DJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFDocument43 pagesDJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFNick_989893No ratings yet
- C & M Bio CytoskeletonDocument40 pagesC & M Bio CytoskeletonA PNo ratings yet
- Cy To SkeletonDocument21 pagesCy To SkeletonJisna JaisonNo ratings yet
- Cytoskeleton Centrosome and CentrioleDocument22 pagesCytoskeleton Centrosome and CentriolekdbuereNo ratings yet
- Cytoskeleton DADocument35 pagesCytoskeleton DAAnonymous NVBCqKWHNo ratings yet
- CytosceletonDocument41 pagesCytosceletonaidar.seralinNo ratings yet
- CytoskeletonDocument65 pagesCytoskeletonSapana SubediNo ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- Muscle ContractionDocument14 pagesMuscle ContractionAlexNo ratings yet
- C YTOSKELETONDocument44 pagesC YTOSKELETONFatma Nadjah MandaNo ratings yet
- Molecular Motors: Ms. M. Banda (B.Pharm, MSC Biochem)Document25 pagesMolecular Motors: Ms. M. Banda (B.Pharm, MSC Biochem)David HamalalaNo ratings yet
- Unit 3 Muscle Physiology - BPT PDFDocument55 pagesUnit 3 Muscle Physiology - BPT PDFRadhikaNo ratings yet
- Lectures 8-10 NotesDocument15 pagesLectures 8-10 Notesapi-3721438No ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- Cytoskeleton (Filament Proteins)Document11 pagesCytoskeleton (Filament Proteins)Siyad AdanNo ratings yet
- Cytoskeleton and Cell Motility 1Document40 pagesCytoskeleton and Cell Motility 1Muhamad Farhan RLNo ratings yet
- Microfilaments. (In Skeletal Muscle Fibers: Kleibsiella Pneumoniae and Shigella, or Yersinia Pestis at 37°CDocument2 pagesMicrofilaments. (In Skeletal Muscle Fibers: Kleibsiella Pneumoniae and Shigella, or Yersinia Pestis at 37°CDavid YapNo ratings yet
- 1 7 CytoskeletonDocument31 pages1 7 CytoskeletonAditya AgrawalNo ratings yet
- Sliding Filament Theory of Muscle Contraction: Muscular TissueDocument8 pagesSliding Filament Theory of Muscle Contraction: Muscular TissueMaryam Nur Azizah RahmahNo ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- MicrofilamentsDocument4 pagesMicrofilamentsSujan maharjanNo ratings yet
- MBB231 Chapter 9 Cytoskeleton Introduction 1197Document28 pagesMBB231 Chapter 9 Cytoskeleton Introduction 1197nathanNo ratings yet
- Skeletal Muscle - 1 Mbbs-1Document30 pagesSkeletal Muscle - 1 Mbbs-1b0t.mc.sundayNo ratings yet
- Enumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonDocument10 pagesEnumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonTathagata ChakravarttyNo ratings yet
- Skeletal Muscle StructureDocument38 pagesSkeletal Muscle Structuretoyy100% (1)
- Cytoskeleton 13.4.20Document39 pagesCytoskeleton 13.4.20NikhilNo ratings yet
- Physiology Slide #1Document18 pagesPhysiology Slide #1anon_974423120No ratings yet
- 2cytology Cytoskel Receptor Transp (3) 1Document43 pages2cytology Cytoskel Receptor Transp (3) 1m.mansoor3377No ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- General Biology Exam 3 Study GuideDocument14 pagesGeneral Biology Exam 3 Study GuideSamantha LapierreNo ratings yet
- Microfilament: in Vitro Self-AssemblyDocument7 pagesMicrofilament: in Vitro Self-AssemblyABHISHEK RATHODNo ratings yet
- The Cytoskeleton: Cell Biology 1st SemesterDocument22 pagesThe Cytoskeleton: Cell Biology 1st SemesterJason CenaNo ratings yet
- Cytology CytoskeletonDocument48 pagesCytology CytoskeletonMitzel Sapalo0% (1)
- UntitledDocument32 pagesUntitleddhwiwdhNo ratings yet
- Cytoskeleton PRESENTATIONDocument36 pagesCytoskeleton PRESENTATIONChaudryNomiNo ratings yet
- Chapter 16 SummaryDocument11 pagesChapter 16 SummaryCharlotteNo ratings yet
- Lecture 3 CytoskeletonfsDocument54 pagesLecture 3 Cytoskeletonfsaidar.seralinNo ratings yet
- Biochemistry of Muscles and Connective TissueDocument41 pagesBiochemistry of Muscles and Connective TissueВіталій Михайлович НечипорукNo ratings yet
- Cell Signaling 5 LessonDocument66 pagesCell Signaling 5 LessonPrathamesh KulkarniNo ratings yet
- 11.sitoskeleton Dan Pergerakan Sel-Dr - IsraDocument40 pages11.sitoskeleton Dan Pergerakan Sel-Dr - IsraTa RaNo ratings yet
- The Cytoskeleton #2: Molecular MotorsDocument34 pagesThe Cytoskeleton #2: Molecular MotorsKimberly ThomasNo ratings yet
- Chapter 9 Actin FloresDocument39 pagesChapter 9 Actin FloresPATRICK MARK FLORESNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- CytoskeletonDocument27 pagesCytoskeletonserenity limNo ratings yet
- Chapter 16 - The Cytoskeleton - 112612Document56 pagesChapter 16 - The Cytoskeleton - 112612Sohan rautNo ratings yet
- Contraction of Skeletal MuscleDocument64 pagesContraction of Skeletal MuscleFhrecy Ann ConsularNo ratings yet
- Cytoskeleton SNDocument3 pagesCytoskeleton SNsamy gNo ratings yet
- Microtubule Based Motor ProteinsDocument6 pagesMicrotubule Based Motor ProteinsIndranil BanerjeeNo ratings yet
- Prepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemDocument37 pagesPrepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemleartaNo ratings yet
- Cellular MovementsDocument27 pagesCellular MovementsngidizzmNo ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- NotesDocument15 pagesNotesGizem OsmanogluNo ratings yet
- Microtubule Oranizing CenterDocument5 pagesMicrotubule Oranizing CenterSasha GutierrezNo ratings yet
- Chapter 2 - NotesDocument22 pagesChapter 2 - NotesanusoumyaNo ratings yet
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Full Chapter Cosmetic Microbiology A Practical Approach 3Rd Edition Philip A Geis PDFDocument53 pagesFull Chapter Cosmetic Microbiology A Practical Approach 3Rd Edition Philip A Geis PDFkarl.lamb140100% (6)
- FAB Classification of Acute Myelogenous Leukemia (AML)Document2 pagesFAB Classification of Acute Myelogenous Leukemia (AML)Christine NazarenoNo ratings yet
- Mitosis in Growing Root Tips: Materials Needed For Each Student PairDocument4 pagesMitosis in Growing Root Tips: Materials Needed For Each Student Pairrajiv pathakNo ratings yet
- A223 P10 Worksheet 032011Document9 pagesA223 P10 Worksheet 032011Dithusha Petchi MuthuNo ratings yet
- Lab 4 DIFFERENTIAL & Special STAINDocument21 pagesLab 4 DIFFERENTIAL & Special STAINAAANo ratings yet
- Cell Injury and Cell Death: Mahmud GhaznawieDocument48 pagesCell Injury and Cell Death: Mahmud GhaznawieLIEBERKHUNNo ratings yet
- Strawberry DNA ExtractionDocument17 pagesStrawberry DNA Extractionabisantiago6131No ratings yet
- DNA Dan ReplikasiDocument41 pagesDNA Dan ReplikasiLollypollybaekbyNo ratings yet
- Biokimia Sistem Endokrin Metabolik 2017Document48 pagesBiokimia Sistem Endokrin Metabolik 2017putuNo ratings yet
- Protien-Synthesis Lesson PlanDocument8 pagesProtien-Synthesis Lesson PlanHAIDEE VASQUEZNo ratings yet
- Biosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future TrendsDocument29 pagesBiosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future Trendsmadhu kammarNo ratings yet
- Advanced Micro :nanotechnologies For Exosome Encapsulation and Targeting in Regenerative MedicineDocument22 pagesAdvanced Micro :nanotechnologies For Exosome Encapsulation and Targeting in Regenerative MedicineSilverStarNo ratings yet
- Lesson Plan Summary Template Feb19-Feb23Document2 pagesLesson Plan Summary Template Feb19-Feb23api-398383485No ratings yet
- Microbiology ReviewerDocument5 pagesMicrobiology ReviewerMarife GuadalupeNo ratings yet
- 1.4 Membrane TransportDocument8 pages1.4 Membrane TransportmariamNo ratings yet
- ScienceDocument142 pagesScienceNhan Tu100% (1)
- Combined HemaDocument28 pagesCombined HemaViena Mae MaglupayNo ratings yet
- 9700 w02 QP 4 PDFDocument12 pages9700 w02 QP 4 PDFY. PurwandariNo ratings yet
- What Is Oxidative Stress 1Document6 pagesWhat Is Oxidative Stress 1Tarmidi MidziNo ratings yet
- Bio 1 SeriesDocument4 pagesBio 1 SerieskisesarevNo ratings yet
- P ET28Document2 pagesP ET28Payal Minocha100% (1)
- Extracellular Matrix Alterations in Metastatic ProcessesDocument18 pagesExtracellular Matrix Alterations in Metastatic ProcessesFernandoNo ratings yet
- Histology of PulpDocument61 pagesHistology of PulpLom SimloteNo ratings yet
- Biosynthesis of Fatty Acids-1Document25 pagesBiosynthesis of Fatty Acids-1Ahmed Imran100% (1)
- L 10 Post Transcriptional ModificationDocument33 pagesL 10 Post Transcriptional ModificationsNo ratings yet
- Chapter Twelve: DNA Replication and Recombination: Comprehension QuestionsDocument13 pagesChapter Twelve: DNA Replication and Recombination: Comprehension QuestionsJeevikaGoyalNo ratings yet
























































































Download as pdf or txt
You might also like
- University of Texas BIO 325 ExamDocument6 pagesUniversity of Texas BIO 325 ExamGeetika InstaNo ratings yet
- Kami Export - Worksheet 14 Central DogmaDocument2 pagesKami Export - Worksheet 14 Central DogmaLindsey TamlinNo ratings yet
- 11.4 Meiosis: Multiple ChoiceDocument5 pages11.4 Meiosis: Multiple ChoiceMING ZHUNo ratings yet
- Amino Acid MetabolsmDocument16 pagesAmino Acid MetabolsmManila MedNo ratings yet
- 17&18) Cell Organization & MovementDocument28 pages17&18) Cell Organization & MovementRNo ratings yet
- BY2201 EmmaC CytoskeletonIandIIDocument62 pagesBY2201 EmmaC CytoskeletonIandIIRebecca ReillyNo ratings yet
- The CytoskeletonDocument10 pagesThe CytoskeletonAhmed khanNo ratings yet
- 2 Cytoskeleton & Cell MotilityDocument47 pages2 Cytoskeleton & Cell MotilityChristineNo ratings yet
- DJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFDocument43 pagesDJ (Microtubules, Microfilaments and Intermediate Filaments) - DJGFNick_989893No ratings yet
- C & M Bio CytoskeletonDocument40 pagesC & M Bio CytoskeletonA PNo ratings yet
- Cy To SkeletonDocument21 pagesCy To SkeletonJisna JaisonNo ratings yet
- Cytoskeleton Centrosome and CentrioleDocument22 pagesCytoskeleton Centrosome and CentriolekdbuereNo ratings yet
- Cytoskeleton DADocument35 pagesCytoskeleton DAAnonymous NVBCqKWHNo ratings yet
- CytosceletonDocument41 pagesCytosceletonaidar.seralinNo ratings yet
- CytoskeletonDocument65 pagesCytoskeletonSapana SubediNo ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- Muscle ContractionDocument14 pagesMuscle ContractionAlexNo ratings yet
- C YTOSKELETONDocument44 pagesC YTOSKELETONFatma Nadjah MandaNo ratings yet
- Molecular Motors: Ms. M. Banda (B.Pharm, MSC Biochem)Document25 pagesMolecular Motors: Ms. M. Banda (B.Pharm, MSC Biochem)David HamalalaNo ratings yet
- Unit 3 Muscle Physiology - BPT PDFDocument55 pagesUnit 3 Muscle Physiology - BPT PDFRadhikaNo ratings yet
- Lectures 8-10 NotesDocument15 pagesLectures 8-10 Notesapi-3721438No ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- Cytoskeleton (Filament Proteins)Document11 pagesCytoskeleton (Filament Proteins)Siyad AdanNo ratings yet
- Cytoskeleton and Cell Motility 1Document40 pagesCytoskeleton and Cell Motility 1Muhamad Farhan RLNo ratings yet
- Microfilaments. (In Skeletal Muscle Fibers: Kleibsiella Pneumoniae and Shigella, or Yersinia Pestis at 37°CDocument2 pagesMicrofilaments. (In Skeletal Muscle Fibers: Kleibsiella Pneumoniae and Shigella, or Yersinia Pestis at 37°CDavid YapNo ratings yet
- 1 7 CytoskeletonDocument31 pages1 7 CytoskeletonAditya AgrawalNo ratings yet
- Sliding Filament Theory of Muscle Contraction: Muscular TissueDocument8 pagesSliding Filament Theory of Muscle Contraction: Muscular TissueMaryam Nur Azizah RahmahNo ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- MicrofilamentsDocument4 pagesMicrofilamentsSujan maharjanNo ratings yet
- MBB231 Chapter 9 Cytoskeleton Introduction 1197Document28 pagesMBB231 Chapter 9 Cytoskeleton Introduction 1197nathanNo ratings yet
- Skeletal Muscle - 1 Mbbs-1Document30 pagesSkeletal Muscle - 1 Mbbs-1b0t.mc.sundayNo ratings yet
- Enumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonDocument10 pagesEnumerating The Structure of Major Protein, Describe The Eukaryotic CytoskeletonTathagata ChakravarttyNo ratings yet
- Skeletal Muscle StructureDocument38 pagesSkeletal Muscle Structuretoyy100% (1)
- Cytoskeleton 13.4.20Document39 pagesCytoskeleton 13.4.20NikhilNo ratings yet
- Physiology Slide #1Document18 pagesPhysiology Slide #1anon_974423120No ratings yet
- 2cytology Cytoskel Receptor Transp (3) 1Document43 pages2cytology Cytoskel Receptor Transp (3) 1m.mansoor3377No ratings yet
- The Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellDocument9 pagesThe Cytoskeleton Is A Network of Filament and Tubules That Extends Throughout A CellHania GulNo ratings yet
- General Biology Exam 3 Study GuideDocument14 pagesGeneral Biology Exam 3 Study GuideSamantha LapierreNo ratings yet
- Microfilament: in Vitro Self-AssemblyDocument7 pagesMicrofilament: in Vitro Self-AssemblyABHISHEK RATHODNo ratings yet
- The Cytoskeleton: Cell Biology 1st SemesterDocument22 pagesThe Cytoskeleton: Cell Biology 1st SemesterJason CenaNo ratings yet
- Cytology CytoskeletonDocument48 pagesCytology CytoskeletonMitzel Sapalo0% (1)
- UntitledDocument32 pagesUntitleddhwiwdhNo ratings yet
- Cytoskeleton PRESENTATIONDocument36 pagesCytoskeleton PRESENTATIONChaudryNomiNo ratings yet
- Chapter 16 SummaryDocument11 pagesChapter 16 SummaryCharlotteNo ratings yet
- Lecture 3 CytoskeletonfsDocument54 pagesLecture 3 Cytoskeletonfsaidar.seralinNo ratings yet
- Biochemistry of Muscles and Connective TissueDocument41 pagesBiochemistry of Muscles and Connective TissueВіталій Михайлович НечипорукNo ratings yet
- Cell Signaling 5 LessonDocument66 pagesCell Signaling 5 LessonPrathamesh KulkarniNo ratings yet
- 11.sitoskeleton Dan Pergerakan Sel-Dr - IsraDocument40 pages11.sitoskeleton Dan Pergerakan Sel-Dr - IsraTa RaNo ratings yet
- The Cytoskeleton #2: Molecular MotorsDocument34 pagesThe Cytoskeleton #2: Molecular MotorsKimberly ThomasNo ratings yet
- Chapter 9 Actin FloresDocument39 pagesChapter 9 Actin FloresPATRICK MARK FLORESNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- CytoskeletonDocument27 pagesCytoskeletonserenity limNo ratings yet
- Chapter 16 - The Cytoskeleton - 112612Document56 pagesChapter 16 - The Cytoskeleton - 112612Sohan rautNo ratings yet
- Contraction of Skeletal MuscleDocument64 pagesContraction of Skeletal MuscleFhrecy Ann ConsularNo ratings yet
- Cytoskeleton SNDocument3 pagesCytoskeleton SNsamy gNo ratings yet
- Microtubule Based Motor ProteinsDocument6 pagesMicrotubule Based Motor ProteinsIndranil BanerjeeNo ratings yet
- Prepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemDocument37 pagesPrepared By:-: Priyanka Yadav M.Sc. Life Sciences Ist SemleartaNo ratings yet
- Cellular MovementsDocument27 pagesCellular MovementsngidizzmNo ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- NotesDocument15 pagesNotesGizem OsmanogluNo ratings yet
- Microtubule Oranizing CenterDocument5 pagesMicrotubule Oranizing CenterSasha GutierrezNo ratings yet
- Chapter 2 - NotesDocument22 pagesChapter 2 - NotesanusoumyaNo ratings yet
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Full Chapter Cosmetic Microbiology A Practical Approach 3Rd Edition Philip A Geis PDFDocument53 pagesFull Chapter Cosmetic Microbiology A Practical Approach 3Rd Edition Philip A Geis PDFkarl.lamb140100% (6)
- FAB Classification of Acute Myelogenous Leukemia (AML)Document2 pagesFAB Classification of Acute Myelogenous Leukemia (AML)Christine NazarenoNo ratings yet
- Mitosis in Growing Root Tips: Materials Needed For Each Student PairDocument4 pagesMitosis in Growing Root Tips: Materials Needed For Each Student Pairrajiv pathakNo ratings yet
- A223 P10 Worksheet 032011Document9 pagesA223 P10 Worksheet 032011Dithusha Petchi MuthuNo ratings yet
- Lab 4 DIFFERENTIAL & Special STAINDocument21 pagesLab 4 DIFFERENTIAL & Special STAINAAANo ratings yet
- Cell Injury and Cell Death: Mahmud GhaznawieDocument48 pagesCell Injury and Cell Death: Mahmud GhaznawieLIEBERKHUNNo ratings yet
- Strawberry DNA ExtractionDocument17 pagesStrawberry DNA Extractionabisantiago6131No ratings yet
- DNA Dan ReplikasiDocument41 pagesDNA Dan ReplikasiLollypollybaekbyNo ratings yet
- Biokimia Sistem Endokrin Metabolik 2017Document48 pagesBiokimia Sistem Endokrin Metabolik 2017putuNo ratings yet
- Protien-Synthesis Lesson PlanDocument8 pagesProtien-Synthesis Lesson PlanHAIDEE VASQUEZNo ratings yet
- Biosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future TrendsDocument29 pagesBiosensors: Microfluidic-Based Droplets For Advanced Regenerative Medicine: Current Challenges and Future Trendsmadhu kammarNo ratings yet
- Advanced Micro :nanotechnologies For Exosome Encapsulation and Targeting in Regenerative MedicineDocument22 pagesAdvanced Micro :nanotechnologies For Exosome Encapsulation and Targeting in Regenerative MedicineSilverStarNo ratings yet
- Lesson Plan Summary Template Feb19-Feb23Document2 pagesLesson Plan Summary Template Feb19-Feb23api-398383485No ratings yet
- Microbiology ReviewerDocument5 pagesMicrobiology ReviewerMarife GuadalupeNo ratings yet
- 1.4 Membrane TransportDocument8 pages1.4 Membrane TransportmariamNo ratings yet
- ScienceDocument142 pagesScienceNhan Tu100% (1)
- Combined HemaDocument28 pagesCombined HemaViena Mae MaglupayNo ratings yet
- 9700 w02 QP 4 PDFDocument12 pages9700 w02 QP 4 PDFY. PurwandariNo ratings yet
- What Is Oxidative Stress 1Document6 pagesWhat Is Oxidative Stress 1Tarmidi MidziNo ratings yet
- Bio 1 SeriesDocument4 pagesBio 1 SerieskisesarevNo ratings yet
- P ET28Document2 pagesP ET28Payal Minocha100% (1)
- Extracellular Matrix Alterations in Metastatic ProcessesDocument18 pagesExtracellular Matrix Alterations in Metastatic ProcessesFernandoNo ratings yet
- Histology of PulpDocument61 pagesHistology of PulpLom SimloteNo ratings yet
- Biosynthesis of Fatty Acids-1Document25 pagesBiosynthesis of Fatty Acids-1Ahmed Imran100% (1)
- L 10 Post Transcriptional ModificationDocument33 pagesL 10 Post Transcriptional ModificationsNo ratings yet
- Chapter Twelve: DNA Replication and Recombination: Comprehension QuestionsDocument13 pagesChapter Twelve: DNA Replication and Recombination: Comprehension QuestionsJeevikaGoyalNo ratings yet